
大豆矮杆片段导入系的外源导入片段分析及基因定位
2020-09-01 13:23魏金鹏赵长江张玉先张海燕任春元于高波
黑龙江八一农垦大学学报 2020年4期
魏金鹏,赵长江,张玉先,张海燕,任春元,于高波
(黑龙江八一农垦大学农学院,大庆163319)
大豆是我国重要的粮食作物之一,含有人类所需的多种氨基酸,特别是亮氨酸、苏氨酸、赖氨酸等,同时还富含多种维生素、不饱和脂肪酸以及微量元素,是人类非常理想的食品营养来源,因此具有非常高的营养价值[1]。除此之外,大豆含有的两种有效成分——大豆异黄酮和大豆皂苷,具有广泛的药理作用和重要的药用价值。因此,大豆的遗传研究一直受到广泛的重视。
作物的株高是重要的农艺性状之一,也是作物研究领域受到广泛关注的性状之一,为数量性状,其表型受到多个基因的共同调控[2]。前人的研究结果显示,大豆株高对产量、荚数、倒伏性等影响较大,同时,株高本身也受到节间长、节数等性状的调节,因此其成为大豆育种工作中育种家重点研究的性状之一[3-4]。在研究过程中人们发现,适当的降低大豆株高,不但能提高植株抗倒伏的能力,还能显著提高大豆的产量[5-6]。早在上个世纪60 年代,美国大豆专家Cooper 教授就提出了矮杆和半矮杆大豆品种的生产种植理论,并利用当时育成的矮杆和半矮杆品种获得了当时的大豆高产纪录[7]。由此可见,对于大豆矮杆相关基因的定位和深入发掘是及其必要的。
目前在遗传育种方面,国内外对大豆株高的研究主要集中在相关QTL 定位和株高与产量的关系方面。张雅娟等[8]以重组自交系群体为研究材料,定位了10 个大豆株高相关的QTL 位点,这些位点分别具有上位性效应和加性效应,可解释55.49%的表型变异。Assefa 等[9-10]利用四个大豆品种,通过全基因组关联对农艺性状进行分析,最终鉴定到了多个与株高和节间数有关的候选位点,部分基因已经在其他植物中被证实参与细胞伸长和植物的生殖生长。高利芳等[11]通过计算分析所收集到的株高相关QTL 信息进行,最终鉴定到了位于5 条染色体上的15 个株高相关的QTL,并筛选出了17 个与株高相关的候选基因。在矮杆基因挖掘方面开展的工作相对较少,Kilen[12-13]利用短节间突变体进行研究分析,其结果显示大豆的矮杆性状由单个基因sb1 所控制,且该基因为隐性基因,而陈恒鹤[14]的研究则显示大豆的矮杆性状是由多个基因共同作用的,包括1 对隐性的主效基因和多对修饰基因,闫昊等[15]的研究结果也证实了这一点,他们通过研究矮杆大豆品种吉密豆1 号与另外两个品种的杂交后代株高数据,分析后发现以一对非完全显性基因为主、多个数量基因为辅共同决定了大豆矮杆性状的表现。另外有报道利用6 个隐性的大豆矮杆突变体进行研究发现,控制矮化的遗传位点分别为df2、df3、df4、df5、df56、df7 和df8,但只有df2 和df5 被定位到了遗传图谱中。任秀慧[16]利用141 份大豆种质资源及3 个重组自交系群体研究大豆矮秆基因,并将相关基因定位在SSR_3 与1248之间;Li 等利用突变体将矮杆基因GmDW1 定位到8号染色体上约460 kb 的区域[17]。截至目前在soybase数据库中,与株高有关的QTL 已经记录了230 个,但与大豆植株矮杆直接相关的QTL 却没有记录,这在一定程度上限制了相关育种工作的进程。
片段导入系(Segments Introgressed Lines,SILs)是指杂交后再经过多次回交选育出来的一套材料,它的基本特征是绝大部分的染色体组成都与轮回亲本完全相同,只在少数几个甚至一个区间内的染色体片段被非轮回亲本(供体亲本)所替代,也称为导入系(Introgression Lines,ILs)或染色体代换系(Chromosome Segment Substitution Lines,CSSLs),含有特定基因时,又叫做近等基因系(Near Isogenic Lines,NILs)[18]。因此,它既能避免其他遗传信息的干扰,又能使整个基因组的QTLs 分解为一个或几个QTLs,对每个QTL 进行单独的分析,从而使主效和微效QTL 都能被挖掘出来,以达到对QTL 的精确定位。片段导入系可以是针对某一特定的基因或性状的导入,也可以非选择性的导入供体亲本的遗传组分,目前第一种导入系应用较多,大多数作物中构建后一种导入系也越来越收到重视。
以导入系为材料研究基因定位已经成为非常有效的分析方式,并且在水稻、玉米、番茄等作物中得到了广泛应用[19-23]。Matus 等[24]利用大麦的染色体导入系鉴定到了一些与麦芽糖含量以及农艺性状相关的基因位点,有趣的是,提供导入片段的供体亲本的相关性状表现却较差;Yamamoto 等[25]通过将籼粳稻杂交后再回交构建导入系,定位到了三个控制水稻抽穗期的QTL;Alpert 和Frary 等[26-27]以番茄为试验材料,通过对栽培种与野生种的杂交再回交,建立了导入群体,并利用该群体发现了与果实重量相关的主效QTL,进一步通过构建图谱进行了精细定位,最终克隆出了该QTL。Casati 等[28]利用不同黄酮含量的玉米植株构建了导入系并研究了紫外线照射下玉米相关基因表达的变化。
研究以携带矮杆基因型S 的片段导入系为研究对象,利用SSR 标记对其进行检测,并对导入片段进行系统分析,对矮杆基因相关的分子标记位点进行了鉴定和分析,研究结果为大豆矮杆相关基因的研究以及矮杆品种的选育奠定了基础。
1 材料与方法
1.1 试验材料
研究的试验材料为7 份片段导入系(SILs),这些导入系均带有矮杆基因S(表1),导入系的轮回亲本为美国早期大豆品种Harosoy,该品种具有抗性强、稳定性好的特点,供体亲本为多个具有优异特性的大豆品种,经6 代或6 代以上的回交培育而成。
1.2 SSR 分析
利用SSR 引物鉴定SILs 中导入片段分布的多态性。PCR 采用20 μL 的反应体系,包括40 ng 基因组DNA,2 μL 10×buffer (含Mg2+),0.4 U Taq 酶,3 μmol 引物和1.5 μL dNTP(2 mmol·L-1),超纯水补齐至20 μL。反应程序为:95 ℃预变性5 min;94 ℃变性30 s,47 ℃退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸5 min。利用聚丙烯酰胺标准测序凝胶对PCR 产物电泳40 min,银染显影后记录标记基因型(带型)。

表1 矮杆片段导入系的系谱Table 1 Information and characters of 7 SILs in experiment
1.3 导入片段分析
对SSR 标记检测结果进行观察分析,以参试材料所显示的条带是否与Harosoy 一致为依据,判断材料中是否含有导入片段。同时利用GGT32 软件构建图示基因型。导入片段长度确定方法参考刘冠明等[29]的方法,SSR 标记位置信息均来自已发表的大豆整合连锁图谱[30]。
1.4 遗传相似性分析
建立SSR 标记检测结果的数据集,根据7 份片段导入系电泳显示的结果,将有迁移条带的标记为1,无迁移条带的记为0,条带缺失的标记为999。利用NTSYS-pc2.1 软件进行相似系数计算及聚类分析,方法采用UPGMA。
1.5 基因定位分析
以SSR 标记检测的结果为基础,若多个片段导入系在同一个标记位点都检测到了染色体片段的导入,则认为该位置存在相关基因或者说该位点与基因紧密关联,用此标记位点来代替基因的位置,以实现对基因的定位。
2 结果与分析
2.1 多态性标记的分析
以Cregan 发表的大豆公共图谱为基础,从中均匀选择分布在20 条染色体上的SSR 标记,最终选择了共计471 个分子标记对全部参试材料进行检测。结果显示,478 个SSR 标记中有66 个标记检测到了多态性(表2)。这些多态性标记分布在除A1 和E 以外的18 条染色体上,每条染色体包含1~8 个多态性标记,平均每条染色体含3.3 个。

表2 SSR 标记在染色体上的分布Table 2 Distribution of SSR markers on chromosomes

续表2 SSR 标记在染色体上的分布Continued table 2 Distribution of SSR markers on chromosomes
2.2 片段导入系与受体亲本的相似性比较
根据SSR 标记电泳后的检测结果可知,7 份SILs 与Harosoy 的相似程度都较高,相似系数为0.956 0~0.979 2(表3),说明所有片段导入系的遗传背景都很一致。
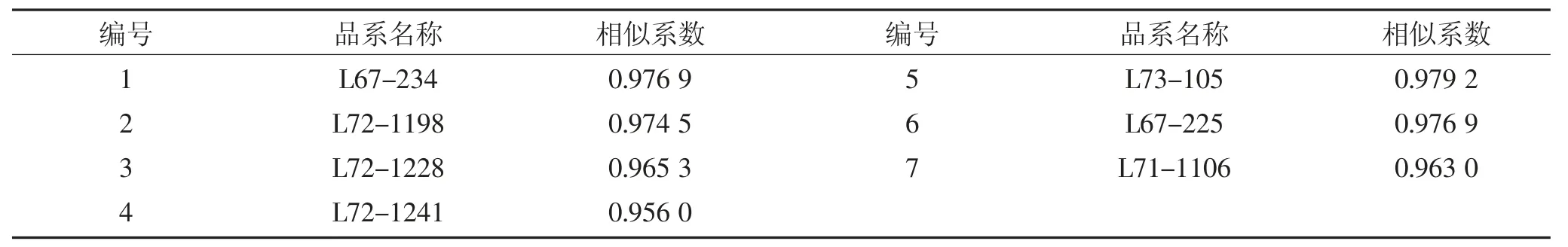
表3 片段导入系与Harosoy 相似系数的比较Table 3 Comparison of similarity coefficient between SILs and recurrent parent
2.3 导入片段分析
利用上面筛选出的66 个SSR 多态性标记,对Harosoy 和矮杆片段导入系进行电泳检测。结果显示,全部标记共检测出了100 个导入片段,其中64个为纯合导入片段,36 个为杂合导入片段。矮杆导入系L71-1106 所含的导入片段数量最多,共有26 个,而L67-234 所含的导入片段最少,仅有7 个,平均每个矮杆片段导入系存在14.3 个片段的导入。矮杆导入系L72-1241 包含最多的纯合导入片段,为18 个,而L67-234 所含的纯合导入片段最少,只有4 个;矮杆导入系L71-1106 含有的杂合导入片段最多,达到20 个,而L72-1241 所含的杂合片段数量最少,只有1 个。全部导入片段的长度为0.17~15.50 cm,平均每个片段的长度为3.63 cm(表4,图1)。

表4 SILs 中导入片段的数量和长度Table 4 Number and length of introgressed segments in SILs

图1 各染色体中导入片段的所在位置Fig.1 Distribution of introgressed segments on chromosomes in SILs
全部导入片段分布在20 条染色体中的18 条上,D1a 和F 染色体上的导入片段都达到了14 个,D1a 染色体上纯合片段和杂合片段分别为6 个和8个,F 染色体上的纯合片段和杂合片段分别为9 个和5 个;G、D2 和D1b 染色体上分别含有13、12 和11个导入片段,A1 和E 两条染色体上没有发现外源片段的导入,剩余染色体上的导入片段数量为1~8 个。
2.4 片段导入系的聚类分析
利用分子标记的检测结果对7 份矮杆片段导入系进行聚类,可以看出,片段导入系间的遗传相似性都呈现出较高的水平。L67-234 与L67-225 相似系数最高,达到0.91,说明他们的遗传背景最为相近;其次相似的依次分别是L72-1198、L72-1228、L72-1241 和L73-105,相似系数分别为0.85、0.80、0.73 和0.71,而L71-1106 与其他材料间的相似系数为0.63,相似程度相对较远(图2)。
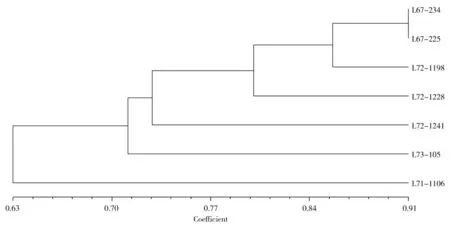
图2 7 份SILs 的聚类分析Fig.2 Clustering analysis of 7 SILs
2.5 矮杆基因S 定位分析
所有的7 份SIL 中都含有矮杆基因S,从材料的系谱中可以看出,所有SIL 的供体亲本中均含有Higan,因此可以认为矮杆基因S 是由供体亲本Higan所提供的。通过对每条染色体上的导入片段进行分析可知,在D1a 染色体的satt383(56.57 cm)位点处,7 份SILs 中有6 份材料都检测到了导入片段,并且都为纯合导入片段,同时在相邻的satt267(57.34 cm)位点处,有4 份SILs 都检测到了杂合导入片段;在D1b 染色体的sat_183(112.63 cm)位点处,有5 份SILs 都发现了纯合的导入片段;在D2 染色体的sat_277(28.8 cm)位点处,有6 份SILs 都发现了纯合的导入片段;而在F 染色体的sct_033(74.13 cm)~satt335(77.70 cm)之间也发现了成簇的导入片段,其中sct_033(74.13 cm)处检测到2 个纯合导入片段,sat_120(75.97 cm)处检测到3 个纯合和1 个杂合导入片段,在satt335(77.70 cm)位点处有3 个SILs 检测到了2 个纯合和1 个杂合导入片段。
因此,推断D1a 染色体上的satt383(56.57 cm)~satt267 (57.34 cm) 区 间、D1b 染 色 体 的sat_183(112.63 cm)位点、D2 染色体的sat_277(28.8 cm)位点以及F 染色体的sct_033(74.13 cm)~satt335(77.70 cm)区间都与矮杆基因S 密切相关,及该基因有很大可能位于这些位点之中。

图3 7 个带有S 基因的SIL 在D1a、D1b、D2 和F 染色体上的导入片段分布Fig.3 Introgressed segments in 7 SILs with S on chromosomes D1a,D1b,D2 and F
3 讨论
将基因片段导入受体亲本是遗传育种中的常见方法之一,对导入片段的有效分析将有利于提高育种效率。研究通过构建图示基因型对SILs 中的导入片段进行分析,共鉴定到100 个导入片段,其中纯合导入片段有64 个,杂合导入片段为36 个,并且导入片段在除A1 和E 之外的18 条染色体上均有分布。对片段导入系的聚类分析说明,研究中的矮杆片段导入系之间的相似性水平较高。L67-234 与L67-225相似系数最高,遗传背景最近,这与他们的选育过程相符,都是仅以Higan 作为供体亲本,再与受体亲本Harosoy 回交6 代选育而成,因此这两份材料所检测到的导入片段也是最少的,分别只有7 个和8 个。相对而言,其他材料的供体亲本除了Higan 之外,都还含有另外一种或一种以上的供体亲本,并且都经历了多次的杂交和回交选育过程,遗传背景更为复杂,因此他们的相似程度略低一些,所检测到的导入片段也就更多,分别为13~26 个,其中L71-1106 检测到的导入片段最多,达到了26 个,这也就解释了该片段导入系虽然供体亲本不是最多的,但其与其他片段导入系的相似系数却是最小的,只有0.63,与其他材料的相似程度相对较远。从片段导入系与轮回亲本的相似性比较以及聚类分析的结果都可以看出,不论是SILs 与轮回亲本Harosoy 还是SILs 之间都存在较高的遗传相似性,说明这套材料具有较为接近的遗传背景,有利于进行基因挖掘方面的研究,定位的结果比较可靠。
在研究中,鉴定到了位于D1a 染色体上的与矮杆基因紧密相关的标记为点satt383(56.57 cm)~satt267(57.34 cm)区间,这与李元龙的研究结果相似,他利用大豆矮化短柄突变体进行研究,并将矮化相关的基因最终定位在到了D1a 染色体上的0.826~1.152 m 处[31],此定位位置与研究结果有所不同,研究在该位置附近仅有1 个SIL 检测到了外源片段的导入,这可能是所用实验材料或标记密度不同所导致。另外,研究结果还发现F 染色体上的sct_033~satt335区间也与矮杆基因紧密相关,有4 个SILs 都检测到了导入片段,结果与袁鹰等[32]的研究结论相似,他们以F2∶3 为研究群体,对控制矮秆性状的隐性基因GmD1 进行分析;结果显示该基因与9 个SSR 标记连 锁, 分 别 是 satt527、satt166、sat_340、satt481、sat_150、sat_245、satt076、satt229 和 satt664。 其 中satt229 位于F 染色体上62.79 cm 处,与sct_033(74.13 cm)和satt335(77.70 cm)的位置都比较接近,这进一步证实了该区域附近确实存在与矮杆性状相关的基因,在下一步的精细定位研究中可重点考虑。李赵博[33]利用具有矮化特性的大豆突变体材料Gmdwarf1 进行研究,结果显示Gmdwarf1 的矮化性状受到一对隐性的细胞核基因所控制,且该基因位于K 染色体上Satt417、Satt264 及Satt337 等3 个分子标记所在的周边区域内,矮化相关基因与Satt337 和Satt417 位点之间的遗传距离都较接近,分别为12.8 cm和14.4 cm。研究中对Satt337 位点也进行了检测,但并未检测到外源导入片段,这可能与所用材料的不同有关。
此外,研究还发现了位于D1b 染色体的sat_183(112.63 cm) 位点和位于D2 染色体的sat_277(28.8 cm)位点都与矮杆基因S 密切相关,而这两个位点在前人的矮杆基因定位研究中并未提及。我们对这两个位点附近的基因分析后发现,与sat_183 相距21 kbp 的基因序列(Glyma.02g256800)编码细胞色素P450 基因家族,该基因家族参与油菜素内酯(BR)和赤霉素(GA)的合成调控,而植物体内BR 和GA 缺失将表现为矮化表型;与sat_183 和sat_277 分别相距25 kbp 和15 kbp 的位置上,各存在一条基因序列(Glyma.02g256200 和Glyma.17g070300)编码富含亮氨酸重复序列的受体蛋白激酶,同时含有丝氨酸/苏氨酸蛋白激酶域,而具有同样功能的BAK1 基因则可以导致植株变矮[34];与sat_277 相距149 bp 的基因序列(Glyma.17g070500)具有编码S-腺苷甲硫氨酸脱羧酶(SAMDC)的功能,该酶参与植物体内多胺的形成,而多胺可调控植物对生长素和细胞分裂素的敏感性,SAMDC 功能缺失将导致植株变得矮小[35]。这些分析结论表明,在sat_183 和sat_277 两个位点附近确实存在产生矮杆性状的基因序列,进一步验证了研究定位结果的可靠性。
研究的结果鉴定到了与前人研究一致的位点,说明了研究结果的可靠性,也证明了充分利用片段导入系进行基因定位研究是可行的,能够进一步检测到由于缺乏多态性而被忽略的标记位点,同时还发现了两处之前未曾报道的新标记位点,这为矮杆基因的发掘研究提供了新的依据,今后可在此基础上进行更加精细的定位,从而准确发掘到矮杆基因,为培育矮杆抗逆高产的大豆新品种奠定基础。
4 结论
在研究的7 份矮杆SILs 中鉴定到了100 个导入片段,其中纯合导入片段有64 个,杂合导入片段为36 个,主要分布在D1a、D1b、D2 和F 染色体上;L67-234 与L67-225 相似系数最高,遗传背景最近,L71-1106 最远;同时也在这四条染色体上发现了与矮杆基因S 紧密关联的标记位点和区间,其中既有与前人研究一致的位点,也有新发现的位点,为进一步研究矮杆基因打下了基础。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
分子催化(2022年1期)2022-11-02
烟草科技(2021年6期)2021-06-24
四川蚕业(2021年4期)2021-03-08
科学之谜(2019年3期)2019-03-28
生物学教学(2018年4期)2018-11-29
电脑知识与技术(2018年19期)2018-11-01
科学之谜(2018年8期)2018-09-29
中学生理科应试(2016年4期)2016-11-19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
