
低温胁迫下不同基因型日本柳杉抗寒性评价
2020-08-31 08:44李庆华王因花燕丽萍刘翠兰吴德军韩邦友李丽
山东农业科学 2020年7期
李庆华,王因花,燕丽萍,刘翠兰,吴德军,韩邦友,李丽
(1.山东省林业科学研究院/山东省林木遗传改良重点实验室,山东 济南 250014;2.山东省调水工程运行维护中心,山东 济南 250014)
日本柳杉[Cryptomeria japonica(L.f.)D.Don]原产于日本,乔木,高达48 m,适合在湿润气候下生长,我国南京、上海、四川、江苏等地均有引种栽培。该树种防风固沙、保持水土,是营造沿海防护林和荒山荒滩造林绿化的先锋树种[1,2]。冻害是北方栽培区种植日本柳杉常见的一种生理性病害,日本柳杉几乎每年都要遭受不同程度的冻害,给其生产带来严重的经济损失。因此,鉴定不同日本柳杉种质的抗寒性,筛选抗寒性强的种质,对日本柳杉产业健康发展、利用创新及其合理栽培区划都具有重要的支撑作用。
目前,关于日本柳杉抗寒性的研究报道甚少。Eguchi等[3]通过对自然条件下的冻害率、细胞渗透浓度等指标的测定比较了日本柳杉的抗寒性,初步筛选出62株抗寒个体,但对各品种抗寒性的了解十分有限。本研究以13个无性系日本柳杉为材料,研究不同低温胁迫处理下其叶片相对电导率及丙二醛、可溶性糖、游离脯氨酸含量等抗寒性指标的变化情况,并利用隶属函数法对其抗寒性进行综合评价,以期为日本柳杉种质的抗寒性评价和抗寒种质选育提供参考。
1 材料与方法
1.1 试验地概况及材料
试验所用日本柳杉(种质编号分别为L1、L2、L3、L4、L5、L6、L7、L8、L9、L10、L11、L12和L13)均种植于日照高科技创新园基地,该地位于东经118°25′~119°39′,北纬35°04′~36°04′,年平均降水量为874 mm,年平均气温为12.6℃,极端最低气温-13.8℃。于2018年8月18日剪取生长整齐健壮的秋梢,培养在装有细沙的盆中,备用。
1.2 试验处理
将秋梢移入人工气候箱内进行不同低温(0、-5、-10、-15℃)胁迫处理,各处理均以5℃/h的速率降温,达到预定温度后保持12 h,以常温(25℃)条件下生长的秋梢作为对照(CK),每处理重复3次,处理完毕后取样进行相关指标的测定。
1.3 测定指标及方法
采用电导法测定相对电导率[4];采用硫代巴比妥酸(TBA)反应法测定丙二醛(MDA)含量,蒽酮比色法测定可溶性糖含量,酸性茚三酮法测定游离脯氨酸(Pro)含量[5]。
1.4 数据分析
采用Microsoft Excel 2019和SPSS 22.0软件进行数据处理和分析。
利用隶属函数法[6]对不同日本柳杉抗寒性进行综合评价。隶属函数值计算公式:
反隶属函数值计算公式:
平均隶属函数值计算公式:
式中,A表示各材料指标的隶属函数值,且A∈[0,1];Hmin、Hmax表示各参试材料的最小值、最大值;B为材料的模糊隶属函数均值,n为指标数。根据上述公式,分别计算出每个种质各项指标的隶属函数值后,用平均隶属函数值进行各种质抗寒能力综合评价,隶属函数均值越大,表明耐寒性越强。
2 结果与分析
2.1 低温胁迫对日本柳杉叶片相对电导率的影响
由表1可以看出,不同日本柳杉叶片相对电导率整体随处理温度的降低而逐渐升高。L4、L6、L9和L11叶片相对电导率随着处理温度的降低变化缓慢,而L3、L5、L8和L12等则表现为迅速增大。25℃处理下,各种质叶片相对电导率差异均不显著(P>0.05),表明此时日本柳杉细胞膜没有受到明显破坏。当低温处理为0℃时,L3、L5、L8和L12叶片相对电导率显著高于其它材料,而其它材料间的叶片相对电导率均差异不显著。当处理温度为-5、-10℃时,不同种质叶片相对电导率存在明显差异,部分达到显著水平。但当处理温度为-15℃时,各种质间叶片相对电导率差异均不显著,表明该温度条件下各种质日本柳杉细胞膜均已遭受严重破坏。
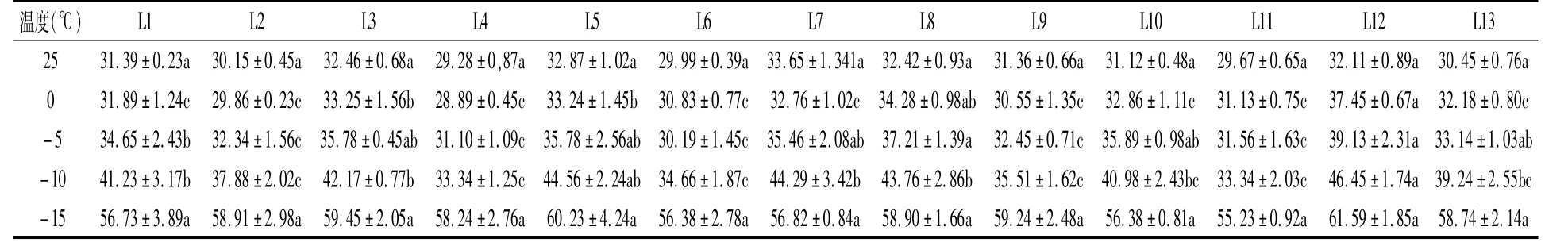
表1 低温胁迫下日本柳杉的叶片相对电导率 (%)
2.2 低温胁迫对日本柳杉叶片丙二醛含量的影响
由图1可以看出,随着处理温度降低,L10叶片丙二醛含量表现为不断升高,其余日本柳杉种质叶片丙二醛含量随处理温度降低整体呈先上升后下降趋势。不同日本柳杉叶片中MDA的最大含量和变化幅度均不同,L5、L13、L12、L8和L3的叶片MDA含量受温度变化影响较大,而L11、L9和L6叶片MDA含量受温度变化影响较小。
2.3 低温胁迫对日本柳杉叶片可溶性糖含量的影响
由图2可以看出,随着处理温度的逐渐降低,13种日本柳杉叶片可溶性糖含量均呈先上升后下降趋势。在25℃处理下,13种日本柳杉叶片可溶性糖含量差异不大;当处理温度低于0℃后,L6叶片可溶性糖含量最高,并在-10℃时达到峰值,为11.24%;其它日本柳杉叶片可溶性糖含量均在-5℃时最高,峰值为4.14% ~9.94%。不同日本柳杉种质可溶性糖含量排序从高到低为L6>L11>L4>L9>L1>L2>L10>L7>L8>L12>L13>L5>L3。
2.4 低温胁迫对日本柳杉叶片游离脯氨酸(Pro)含量的影响
由图3可以看出,13种日本柳杉叶片Pro含量均随处理温度的降低呈先上升后下降的趋势。L2、L4、L6、L9和L11叶片Pro含量的最高值均出现在-10℃时,其中,L11最高,L6、L4次之,L9较低,L2最低,但均分别比25℃时增加5.49、5.44、5.30、3.86倍和2.40倍。L1、L3、L5、L7、L8、L10、L12和L13叶片Pro含量峰值均出现在-5℃时,含量从高到低依次为L1>L10>L13>L3>L8>L7>L5>L12,分别比25℃时增加2.72、2.64、2.62、2.09、1.86、1.82、1.82倍和1.60倍。
2.5 13个日本柳杉抗寒性综合评价
为了较全面地研究本试验材料的抗寒性,运用隶属函数法,通过4个抗寒指标平均隶属度进行综合评价,确定不同日本柳杉种质的抗寒性。由表2可以看出,13个日本柳杉抗寒性从强到弱依次为:L11>L6>L4>L9>L2>L1>L10>L7>L13>L8>L3>L5>L12。

表2 日本柳杉叶片抗寒性评价
3 讨论与结论
低温胁迫下,植物会发生一系列的生理生化和大分子变化以及生物膜结构改变等以适应低温逆境[7,8]。相对电导率和丙二醛含量的变化能够直观反映抗逆环境中植物细胞膜的受损程度。大多数研究表明,植物在低温逆境条件下细胞膜透性增大、胞内电解质外渗,进而引起电导率升高[5]。本研究中,日本柳杉在低温胁迫处理下电导率和MDA 含量均增大。该结果与其它物种[9,10]上的研究报道类似。
可溶性糖和游离脯氨酸是植物体内的重要渗透调节物质,其含量高低与植物耐寒性密切相关[11,12]。在低温胁迫下,糖类代谢主要调节植物体内生物合成和蛋白质分解之间的平衡,降低冰点、缓和细胞质过度脱水,从而调节植物在逆境胁迫下的适应性,提高植物的抗寒能力[13,14]。植物在低温逆境中可通过调节原生质与环境的渗透平衡,防止细胞结构受破坏,增强保护作用[15]。本研究表明,日本柳杉可溶性糖和游离脯氨酸含量均随处理温度的降低呈先上升后下降的趋势。这与李瑞雪[16]、刘贝贝[17]等的研究结果一致。
采用单一指标评价参试材料的抗寒性会产生较大误差,这可能由于样品本身、自然环境和生理状况的不同,导致不同的生理指标对同一批次样品的抗寒性评价结果不一致[18]。采用隶属函数法能够较全面地分析判断植物抗寒能力,尽可能地克服采用单个指标的片面性[19]。目前这种方法已经在板栗、果树和扁桃等树种的抗寒性鉴定中广泛应用[18,20,21]。本研究运用隶属函数法计算了与日本柳杉叶片抗寒性密切相关的4个指标的平均隶属度,对13个日本柳杉叶片抗寒性进行了综合分析排序,并初步筛选得到高抗寒品种。说明采用隶属函数法能够较为科学地评价和区分日本柳杉的抗寒性强弱,具有较强的适用性。本研究对拟引入的日本柳杉株系在本地的抗寒表现进行科学预判,以避免盲目引种带来的生产损失。
本研究利用隶属函数法综合评价13个日本柳杉抗寒性,得出其抗寒性由强到弱依次为:L11>L6>L4>L9>L2>L1>L10>L7>L13>L8>L3>L5>L12,并筛选出L11、L6、L4和L9共4个耐寒性强的种质。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
热带作物学报(2022年5期)2022-06-01
干旱地区农业研究(2022年3期)2022-05-24
中国种业(2022年1期)2022-01-27
矿冶工程(2021年4期)2021-09-15
山西农业科学(2020年10期)2020-10-16
世界有色金属(2020年15期)2020-10-10
四川蚕业(2020年4期)2020-02-10
园艺与种苗(2015年10期)2015-02-27
