
5个郁金香品种日光温室内光合特性的比较研究
2020-08-31 08:44乔谦王雪苏亚静王江勇陶吉寒孙忠奎
山东农业科学 2020年7期
乔谦,王雪,苏亚静,王江勇,陶吉寒,孙忠奎
(1.山东省果树研究所,山东 泰安 271000;2.泰安时代园林科技开发有限公司,山东 泰安 271000;3.山东农业大学林学院,山东 泰安 271018)
郁金香(Tulipa gesnerianaL.)属百合科郁金香属多年生鳞茎草本植物,别名洋荷花、草麝香,具有很高的观赏价值和经济价值,是世界公认的名贵花卉,被誉为“花卉王国中的皇后”[1-3],国际市场上供不应求。据统计,全世界约有40~150种野生郁金香,栽培品种达1万多个,其中约150个为大量生产品种[4,5]。我国共有野生郁金香属植物16种,集中分布于新疆北部、内蒙古西部及中东部省区,但自主栽培品种较少,多受限于我国气候条件、栽培技术等因素,此外,种球主要依赖进口,且退化严重,需年年种植[2]。近年来,随着各大城市的广泛栽培,郁金香种苗供给更是形成了巨大的市场缺口。
为满足元旦、春节等节日的用花需求,国内郁金香栽培多采用日光温室促成栽培[5-8]。日光温室栽培过程中易出现光照不足、空气流通不畅的情况,影响植物光合能力[9]。而光合能力是产量和品质形成的基础,光合太弱会导致植物生长缓慢、生物量积累少,进而影响开花结果等生理特性。所以探索合适的栽培条件并筛选光合能力较强的品种尤为重要。本试验研究日光温室栽培条件下不同郁金香品种的光合参数日变化、环境因子和光响应曲线,并分析环境因子间及光合参数与环境因子的相关性,以期为郁金香的设施栽培提供参考。
1 材料与方法
1.1 材料
供试郁金香品种为‘检阅’(Parade)‘大王子’(Ad Rem)‘小王子’(World’s Favourite)‘金检阅’(Golden Parade)和‘卡罗拉’(Carola),混合种植于山东省果树研究所金牛山基地日光温室大棚内。
1.2 试验方法
每品种随机选择3株长势健壮、无病虫害且生长一致的蕾前植株。每株选取外轮第2~3片健康成熟功能叶作为测试叶,测量位点为叶片最宽处[2],测试前擦净叶片表面灰尘。
1.2.1 环境因子和光合参数日变化的测量 于2018年1月25日(晴天)9∶00—17∶00,使用CIRAS-3便携式植物光合作用测定仪,每隔2 h测定1次净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、水分利用效率(WUE)等光合参数[10]。同时记录温室内相应环境因子变化,包括光合有效辐射(PAR)、气温(Ta)、叶片温度(Tl)、大气CO2浓度(Ca)、水汽压(e)和空气相对湿度(RH),并计算气孔限制值(Ls)。计算公式:
1.2.2 光合响应曲线制作 于晴天上午9∶00—11∶00,采用CIRAS-3便携式植物光合作用测定仪,选用LED红/蓝光源,气路开放,分别测定光合有效辐射梯度为0、50、100、150、200、400、600、800、1 000、1 200、1 600、2 000μmol/(m2·s)下的Pn值。整理数据绘制Pn-PAR响应曲线,通过直角双曲线的修正模型拟合曲线计算表观量子效率(AQY)、光饱和点(LSP)和光补偿点(LCP)[11,12]。
1.3 数据处理
采用Microsoft Excel 2010软件进行数据统计分析和作图,用SPSS 24.0软件进行方差分析和相关性分析。
2 结果与分析
2.1 不同郁金香品种叶片环境因子日变化及其相关性分析
9∶00—17∶00,日光温室内不同郁金香品种叶片环境中PAR变化均呈先上升后下降趋势。其中,‘检阅’和‘小王子’均在11∶00达到峰值,而‘大王子’‘金检阅’和‘卡罗拉’则在11∶00后持续增加,于13∶00达到峰值(图1A)。
日光温室内气温整体呈先升高后降低趋势。其中,‘检阅’‘大王子’和‘金检阅’13∶00达到最大值,‘小王子’在15∶00达到最大值(27.66℃),而‘卡罗拉’则在11∶00达到最大值,仅为27.12℃(图1B)。
不同郁金香品种的叶片温度与气温变化趋势相同,均为单峰变化曲线(图1C)。
日光温室内大气CO2浓度9∶00最高,急剧下降至11∶00后缓慢下降,15∶00—17∶00略有上升,日均值分别为474.65(‘检阅’)、465.58(‘大王子’)、460.97(‘小王子’)、452.53(‘金检阅’)、450.36(‘卡罗拉’)μmol/(m2·s),见图1D,品种间差异不显著(P>0.05)。
水汽压(e)变化与PAR一致,且各品种间变化趋势一致,均呈先升高后降低再缓慢升高的趋势,11∶00或13∶00达到最大值(图1E)。
空气相对湿度(RH)变化趋势如图1F所示,除‘小王子’呈现先升高后降低趋势,‘卡罗拉’和‘大王子’均从9∶00开始下降直至15∶00出现升高趋势,其它品种则呈逐渐下降趋势。
由表1可以看出,Ta和Tl相关系数为0.998,呈极显著正相关,说明叶片温度主要受气温影响。e与PAR相关系数为0.964,呈极显著正相关。其它环境参数间的相关性均未达显著水平。
2.2 不同郁金香品种光合参数日变化分析
由图2A可以看出,不同郁金香品种净光合速率的日变化趋势一致,均呈双峰型,出现“光合午休”现象,峰值分别出现在11∶00、15∶00。其中,‘检阅’的最高峰值[7.66μmol/(m2·s)]和日平均值[2.07μmol/(m2·s)]均最大,‘小王子’‘大王子’次之,‘金检阅’和‘卡罗拉’较小。
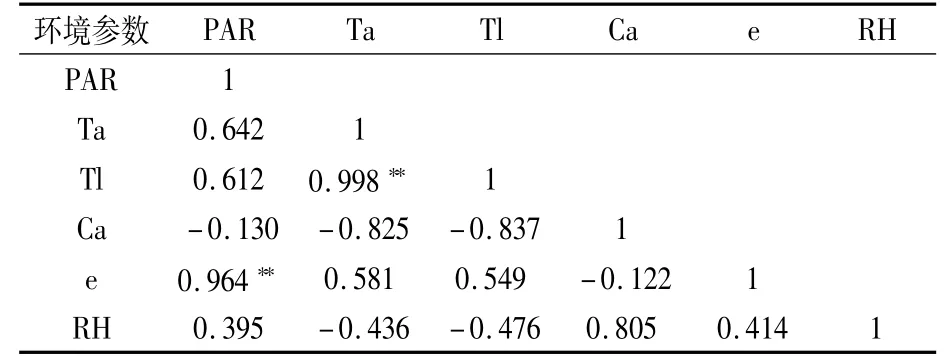
表1 日光温室内叶片环境因子相关性分析
不同郁金香品种气孔导度日变化中(图2B),‘大王子’‘小王子’和‘金检阅’均呈先升高再下降后逐渐平缓的趋势,11∶00达到峰值,而‘卡罗拉’则一直表现稳定,9∶00到17∶00变化不大,‘检阅’则在9∶00到13∶00比较稳定,13∶00后开始下降。不同品种Gs峰值和日均值以‘大王子’最大,分别为916.00、541.15 mmol/(m2·s)。
不同品种胞间CO2浓度的日变化趋势大体一致(图2C),呈现早晚高中午低的双谷型变化曲线,从9∶00开始快速下降,11∶00到15∶00先上升后下降,15∶00后逐步上升;最高值均出现在9∶00,其中‘检阅’的胞间CO2浓度最大,为594.00 μmol/mol,其次是‘大王子’(554.20μmol/mol)和‘小王子’(527.00μmol/mol)。
不同品种蒸腾速率日变化有单峰型和双峰型两种(图2D),‘小王子’为明显的双峰型,分别在11∶00和15∶00达到峰值,其它品种均为单峰型,‘检阅’在13∶00达到峰值,‘大王子’‘金检阅’和‘卡罗拉’则在11∶00达到峰值,之后缓慢持续下降。‘检阅’的蒸腾速率峰值最高,为7.13 mmol/(m2·s),‘大王子’的日均值最大,为4.73 mmol/(m2·s)。
水分利用效率的日变化如图2E所示,‘检阅’呈单峰型曲线变化,其它品种均呈双峰型变化,最高值均出现在11∶00。‘检阅’的最高值高于其它品种,为1.52μmol/mmol;而‘小王子’的日均值最大,为0.41μmol/mmol。
由图2F可以看出,气孔限制值日变化趋势与WUE具有高度相似性,‘检阅’和‘小王子’为单峰型,峰值分别出现在11∶00(0.106)和15∶00(0.064),其它品种均呈双峰型,峰值分别出现在11∶00和15∶00。
2.3 不同郁金香品种光合参数与环境因子日变化的相关性分析
由表2可以看出,不同郁金香品种的Pn均与PAR、e、RH的相关性较高且均呈正相关,说明适当提高温室内光照和湿度有利于郁金香光合速率的提高,进而加快营养生长。
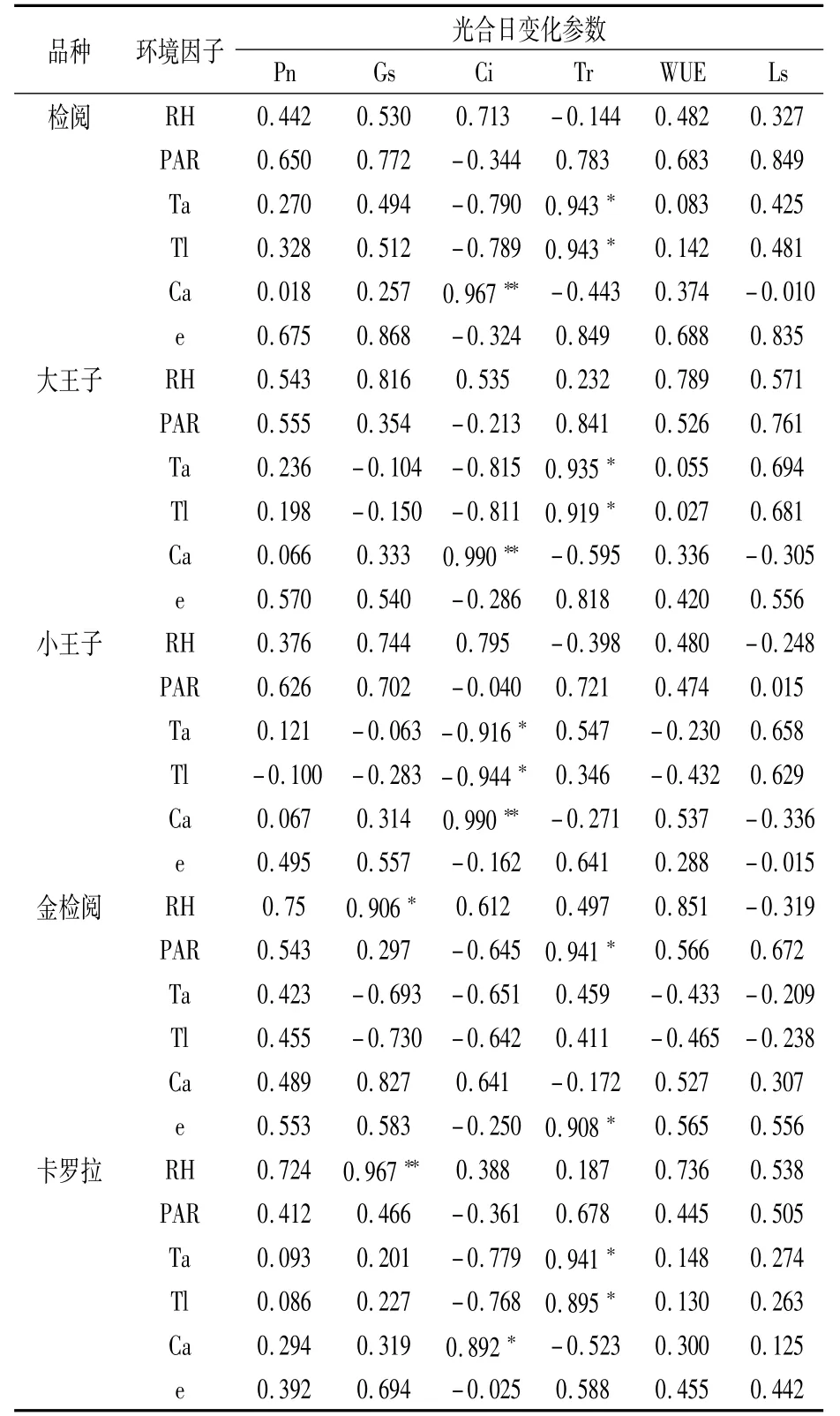
表2 不同郁金香品种光合参数与环境因子日变化的相关系数
Gs也与PAR、e、RH呈较高正相关性,其中‘金检阅’和‘卡罗拉’的Gs分别与RH呈显著、极显著正相关,相关系数分别为0.906、0.967,表明相对湿度是影响气孔导度的重要因素。
不同品种的Ci均与Ca正相关,‘检阅’‘大王子’‘小王子’的相关性达极显著水平,‘卡罗拉’达显著水平;‘小王子’Ci还与Ta、Tl呈显著负相关,相关系数分别为-0.916、-0.944,表明温度也是影响胞间CO2浓度的重要因素。
‘检阅’‘大王子’和‘卡罗拉’的Tr主要与Ta、Tl有关,呈显著正相关,‘金检阅’的Tr则与PAR、e呈显著正相关。
不同品种的WUE、Ls与各环境参数均未表现出显著性相关。其中,水分利用效率主要与PAR、RH呈正相关;气孔限制值主要与PAR呈正相关,部分品种还与温度或湿度呈正相关,但均与CO2浓度未体现明显相关性。因此,温室内通过补充光照、提高温度和湿度可达到提升光合速率的目的。
2.4 不同郁金香品种的光响应曲线
由图3可以看出,不同郁金香品种的光响应曲线变化趋势基本一致,具体表现为,当光合有效辐射在0~600μmol/(m2·s)时,净光合速率随光合有效辐射的升高而快速增加,当光合有效辐射在800 μmol/(m2·s)以上时,净光合速率缓慢增加直至稳定状态。‘检阅’各光合有效辐射对应的净光合速率均大于其它品种,最高可达26.9μmol/(m2·s),‘大王子’‘小王子’和‘卡罗拉’的最大光合速率相当,‘金检阅’最小,为21.8μmol/(m2·s)。
通过对不同品种光响应曲线的拟合曲线方程可得表3。不同郁金香品种的光饱和点从大到小依次为‘检阅’>‘大王子’>‘金检阅’>‘小王子’>‘卡罗拉’,平均值为717.21μmol/(m2·s);光补偿点从大到小依次为‘大王子’>‘金检阅’>‘卡罗拉’>‘小王子’>‘检阅’,平均值为56.80 μmol/(m2·s);表观量子效率以‘检阅’最高,为0.054,‘卡罗拉’次之,其它品种均为0.043,平均值为0.045。
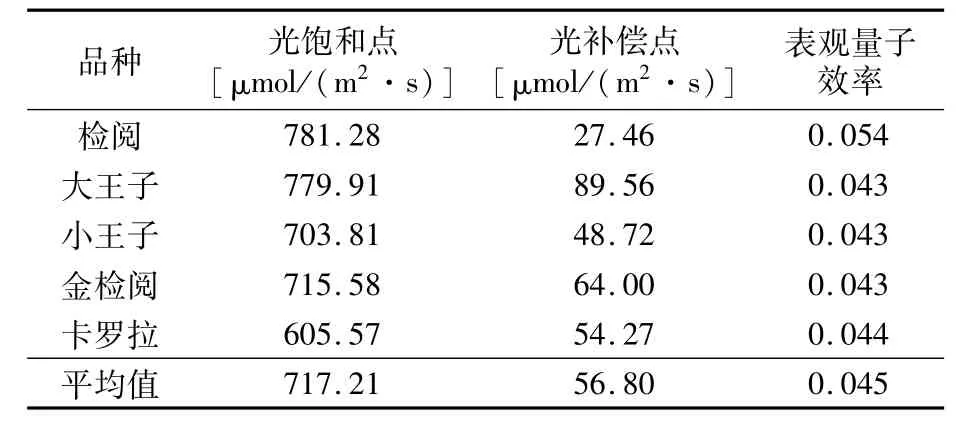
表3 不同郁金香品种光合-光响应参数
3 讨论与结论
植物叶片光合作用日变化是植物物质积累与生理代谢的基本过程,也是分析环境因素影响植物生长和代谢的重要手段[13,14]。植物光合作用日变化通常有两种典型方式,即双峰型和单峰型[15]。姜文正等[2]认为室外栽培‘上农早霞’郁金香的净光合速率日变化呈双峰型。本研究中,不同郁金香品种的净光合速率日变化结果与其一致,但温室内最高光合有效辐射仅为287.40 μmol/(m2·s),远低于光饱和点对应的光合有效辐射[781.28μmol/(m2·s)],推测这种“光合午休”现象是温度、湿度、大气CO2浓度等环境因子综合作用的结果。该结果与日光温室栽培百合、草莓等园艺作物一致[10,16]。此外,不同郁金香品种的净光合速率表现为‘检阅’>‘小王子’>‘大王子’>‘金检阅’>‘卡罗拉’,‘检阅’的日均值最大,但仅为2.07μmol/(m2·s),并且‘金检阅’和‘卡罗拉’的日均值均为负值,表明现温室环境无法满足郁金香的生长需求,急需改善温室条件。通过分析其它光合指标,发现蒸腾速率[最高7.13 mmol/(m2·s)]明显高于草莓[16]、甜椒[17]、切花菊[18]、百合[10]等园艺作物,而水分利用效率又明显低,最高仅为1.52μmol/mmol,大部分为负值,这符合Robredo等[19]得出的气孔导度对植物获取CO2和水分散失的调节中无法实现优化的条件;并且气孔限制值也整体偏小,最大仅为0.106。因此建议应结合温室内水分条件,补充水分使水汽压提升至1.0~1.1 hPa、温室内相对湿度增至48%~53%。
本试验中,通过对光合日变化特征参数与环境因子相关性进行分析发现,温度与多数郁金香品种的蒸腾速率呈显著正相关,同时与胞间CO2浓度具有较强的负相关,进而影响了净光合速率。相关研究也表明温度过高或过低均降低净光合速率[20-22],结合本试验温室内温度变化认为需降低午间温度,适当提高早晚温度,使白天温度达到24~27℃,从而提高光合速率、降低蒸腾速率,进而促进郁金香提早开花赶上元旦、春节等节日。
不同郁金香品种的LCP平均值为56.80 μmol/(m2·s),LSP值平均为717.21μmol/(m2·s)。其中,LCP稍高于前人研究结果[22,23],LSP较低,但净光合速率却较高。表明这5个郁金香品种均能够高效利用光能。本试验中温室光照最高仅为287.40μmol/(m2·s),远低于饱和光强,导致净光合速率明显低下,部分时间段出现负值,限制了郁金香生长。此问题在其它温室栽培园艺作物中也普遍存在,如百合[10]、切花菊[18]、甜椒[17]、黄瓜[24]、番茄[25]等。因此,冬季日光温室栽培条件下,应适当增加光照强度,增强光合作用,促进植物有机物积累。
植物利用弱光能力的重要指标包括LCP和AQY,植物利用强光的能力则是通过LSP反映[13,26]。5个郁金香品种中,‘检阅’具有最低的LCP和最高的LSP、AQY,说明其既能利用弱光也能充分利用强光,对光照条件的适应范围更广,而‘大王子’具有最高的LCP和较高的LSP,表明其吸收利用弱光能力差,更加适应强光环境;5个品种生长情况也表明‘检阅’长势快于其它品种。因此,本研究认为‘检阅’更适宜在日光温室内栽培。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
中老年保健(2021年4期)2021-08-22
农业与技术(2020年12期)2020-07-04
时代英语·高一(2019年5期)2019-09-03
车迷(2019年10期)2019-06-24
车迷(2018年11期)2018-08-30
新作文·小学高年级版(2017年10期)2017-11-13
新课程·下旬(2017年7期)2017-08-14
小猕猴智力画刊(2017年5期)2017-05-25
好孩子画报(2014年11期)2014-11-12
