
白桦BpPR1基因生物信息学分析
2020-08-28 11:20安玉婷李然红陈鑫刘丹
湖北农业科学 2020年12期
安玉婷 李然红 陈鑫 刘丹


摘要:利用生物信息学方法,对白桦BpPRI理化性质、亲水/疏水性、跨膜结构、二级结构、三级结构及与其他植物PRI蛋白的同源性进行了预测,同时对该基因启动子的顺式作用元件以及基因结构进行了预测和分析。结果表明,①BpPR1的cDNA全长为816 bp,CDS为53lbp,编码由176个氨基酸构成的稳定亲水性蛋白,存在跨膜运输结构,属于CAP蛋白家族的一员。白桦BpPR1与葡萄、枣亲缘关系较近;②BpPR1基因组结构中含有2个外显子和3个内含子;③BpPR1启动子含有多个真核生物启动子的基本元件CAAT-box和TATA-box,还含有与低温响应、脱落酸响应、茉莉酸甲酯响应、生长素响应、水杨酸响应、细胞分裂素响应相关的响应元件以及MYBHvl结合位点等,说明该蛋白可能受多种生长调节剂调控,参与生物与非生物胁迫过程。
关键词:白桦(Betula platyphylla);病程相关蛋白;生物信息学
中图分类号:Q789
文献标识码:A
文章编号:0439-8114( 2020)12-0171-04
DOI:10.1408 8/j .cnki.issn0439-8114.2020.12.038
开放科学(资源服务)标识码(OSID):
白桦(Betula platyphylla)为桦木科桦木属落叶木本植物,在亚欧大陆中分布较多。而在中国,其主要分布于东北、华北及西北等地区[1]。白桦具有适应能力强、生长快、材质优等特点,已成为当今社会最有应用价值及发展潜力的树木种类之一,对白桦相关研究也越来越受到重视[2]。同时,白桦形态优美、表面光洁,常作为城市绿化树种[3]。白桦皮及白桦汁具有很高的药用价值,近年来,白桦已被列入重点的科技研究项目之一[4]。植物病程相关蛋白(Pathogenesis—related protein,PR)是植物在生物或非生物胁迫下产生的一类抵御不良环境的蛋白总称。PR蛋白最早在烟草中被发现,在病毒侵染植株后产生,并且在侵染及未侵染部位PR基因都有大量表达[5]。PR广泛存在于动物、植物中,在单子叶和双子叶植物中存在更为广泛,由17个蛋白家族组成,其中,已被发现功能的有PR2(β一1,3一葡聚糖酶)、PR3(几丁质酶)、PR5(类甜蛋白)、PR6(蛋白酶抑制剂)、PR7(蛋白内切酶)、PR9(过氧化物酶)、PRlO(类核糖核酸酶)、PR12(防卫素)、PR13(硫素)、PR14(脂质转运蛋白)等。当植物受到病原体感染时,PR的表达量显著提高[6-8}。PR1作为病程相关蛋白家族成员之一,现已经被证实具有抵抗真菌入侵、限制病毒扩散及抵御逆境胁迫能力等功能。白桦类病斑突变体Imd对半知菌亚门的链格孢菌[ Alternaria alternata( Fr.) Keissler]具有显著抗性,BpPR1基因表达量在病原菌入侵时显著提高。[9]。
本研究参考白桦转录组和基因组数据[10],获得白桦BpPR1基因的序列信息,利用生物信息学方法对其理化性质、疏水性/亲水性、蛋白结构功能域、二级结构、三级结构、蛋白质修饰位点及同源性等进行了预测,对其基因结构进行分析,并与其他植物PR1基因结构进行了比较,为进一步研究该基因的功能及其表达调控机制提供参考。
1 材料与方法
1.1 材料来源
白桦BpPR1基因启动子和基因序列从CoCe网站获得( https://genomevolution.org/coge/)。擬南芥(Arabidopsis thaliana)、葡萄(Vitis vinifera)、核桃(Juglans regia)和毛果杨(Populus trichocarpa)的基因结构信息从CoGe网站获得。葡萄、枣(Ziziphusjujuba)、榴莲(Durio zibethinus)、毛白杨(PoprLILLs to-mentosa)、橡胶树(Hevea brasiliensis)、耧斗菜(Aquile-gia viridiflora)、豇豆(Vigna unguiculata)、菜豆(Phaseolus vulgaris)、核桃(Juglans regia)、可可(Theobroma cacao)、鼠尾草(Herrania umbratica)、红车轴草(Trifolium pratense)、花芽龙眼(Dimocarpus longan)、克莱门柚(Citrus clementina)、毛果杨(Popu-lus trichocarpa),木豆(Cajanus cajan)、栓皮栎(Quer-cus suber)等植物基因信息在NCBI( https://www.ncbi.nlm.nih.gov/)数据库中获得。
1.2 方法
本研究中用到的软件及网址见表1。
2 结果与分析
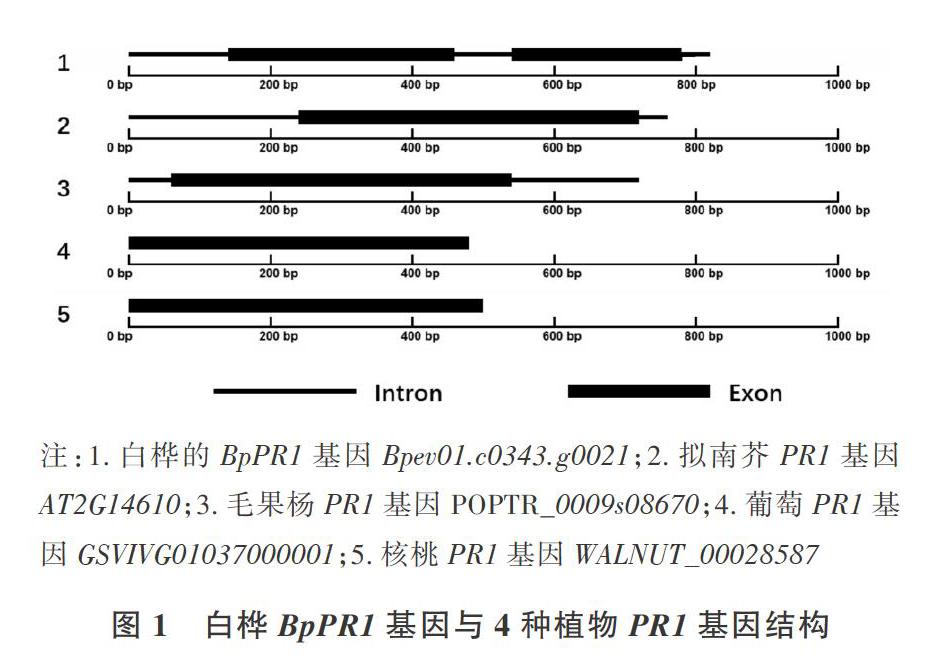
2.1 白桦BpPR1基因结构分析
利用在线软件GSDS(http://gsds. cbi. pku. edu.cn)[11]对BpPR1基因结构进行分析,并与拟南芥、葡萄、毛果杨、核桃等植物的PR1基因结构进行比较。结果(图1)表明,BpPR1基因中有3个内含子和2个外显子,CDS为531 bp;毛果杨、拟南芥PR1基因中都含有一个外显子和两个内含子,CDS长度为485 bp;葡萄PR1基因的CDS也为485 bp,但没有内含子;核桃PR1基因没有内含子,CDS为503 bp;拟南芥、葡萄、毛果杨PR1基因的CDS都是485 bp,其基因结构的差别在于内含子不同。
2.2 白桦BpPR1理化性质预测
利用ProtParam软件对白桦BpPRI的氨基酸序列进行分析,结果表明,BpPR1基因cDNA全长为816 bp,基因编码区(CDS)为cDNA序列中第37至661个碱基,共编码176个氨基酸,分子质量19 265.90D,理论等电点为8.93,负电荷残基总数(Asp+Clu)为10,正电荷残基总数( Arg+Lys)为15,分子式为C842:H1312 N246,O250,S12,总原子数2 662,不稳定指数(Instability index,Ⅱ)为31.7,脂肪系数(Aliphatic in-dex)为1.36,平均亲水性(GRAVY)水平为-0.178,说明该蛋白为稳定的亲水性蛋白。该蛋白中含量最多的氨基酸为缬氨酸,占1 1.4%,其次为甘氨酸,占9.7%,含量最少的为谷氨酸和苯丙氨酸,分别为1.7%和1.1%。通过ProtScale软件分析预测,该蛋白质整个肽链中均含有亲水性和疏水性氨基酸,得分最大值为3.311,在第11个氨基酸处,最小值为-2.656,在第265个氨基酸处,氨基酸的负值分值大,根据蛋白质亲水和疏水的得分判定该蛋白为亲水性蛋白(图2)。
2.3 BpPR1信号肽和跨膜结构分析
通过SignaIP-4.1预测BpPRI信号肽及跨膜结构,结果(图3)表明,原始剪切位点的分值(C值)在第25个氨基酸时有最大值,为0.445,信号肽的分值(S值)在第14氨基酸时有最大值,为0.990,综合剪切位点的分值(Y值)在第25个氨基酸时有最大值,为0.644,其D-cut off为0.717 06(>0.5),表明BpPRI蛋白含有信号肽,存在跨膜运输结构。
2.4 BpPR1二级、三级结构预测
蛋白质二级结构是指肽链由氢键构成有规则的卷曲折叠具有周期性的空间结构,使用SOPMA对白桦BpPRI二级结构进行分析预测。结果表明,该蛋白9.83%可能会形成螺旋(Helix),27.12%可能形成延伸链( Strand),4.14%可能形成转角(Turn),58.64%为无规卷曲(Random coil),无二硫键的形成,没有特殊的二级结构。用SWISS-MODEL对BpPR1蛋白进行三级结构的预测(图4),不同类型二级结构进一步折叠,形成具有空穴的高级结构,可能是某些互作物质的结合位点。
2.5 氨基酸序列同源性对比
在NCBI数据库中分别搜索葡萄、枣、榴莲、毛果杨、榴莲、橡胶树、耧斗菜、豇豆、菜豆、核桃、可可、鼠尾草、红车轴草、花芽龙眼、克莱门柚、毛果杨、木豆、栓皮栎等17种植物的PRI序列,与白桦BpPRI氨基酸序列进行同源性分析。结果(图5)表明,病程相关蛋白BpPRI与葡萄、枣亲缘关系较近,而与榴莲、毛白杨等植物亲缘关系较远。
2.6 白桦BpPR1基因启动子序列分析
利用PlantCare对白桦BpPR1基因启动子顺式调控元件进行分析。结果(表2)表明,BpPR1基因含有多个真核生物启动子的基本元件CAAT-box和TATA-box,含有茉莉酸甲酯响应的顺式作用元件、光响应顺式作用元件、低温响应的顺式反应原件、脱落酸响应的顺式作用原件、MYBHvl结合位点、生长素响应的顺式作用元件、水杨酸响应的顺式作用元件、细胞分裂素的顺式作用元件等,说明PR1基因与逆境胁迫密切相关。乙烯(ET)、茉莉酸(JA)、水杨酸(SA)是诱导产生抗病性重要的信号分子,可诱导植物中PRI的表达[12,13]。白桦BpPR1基因与多种植物生长调节剂响应有关。
3 小结与讨论
病程相关蛋白是植物中一类重要的蛋白,植物在受到病原体侵染时,水杨酸( Salicylicacid,SA)、茉莉酸( Jasmonicacid,JA)、乙烯(Ethylene,ET)等生长调节剂含量会随之改变,同时PR基因会选择性表达‘14.15]。PR1作为病程相关基因重要的一员,对其结构功能的研究也在逐步进行。侯丽霞等[16]研究表明葡萄PRI可对多种胁迫产生响应应答,霜霉病菌、低温、盐、干旱、水杨酸、脱落酸、茉莉酸、一氧化氮、过氧化氢和硫化氢等可诱导其大量表达。马立功等[17]对向日葵的研究发现PR1在盐胁迫、干旱胁迫、核盘菌及其代谢产物的诱导下可上调表达,将该基因转接到烟草中,获得转基因烟草,转基因植株也提高了对核盘菌的抗性。PR1基因的表达受多种基因调控,PR1基因是否具有抗病性与其基因结构有关。张凯敏等[18]研究发现刚毛柽柳的TbPR1基因的编码区长为537 bp,共编码178个氨基酸,与白桦BpPR1较为接近。Li等[19]研究发现葡萄中烟草野火病菌只有在碱性PRI存在的情况下才不表达;Bonasera等[20]将PR1基因转入到苹果中,发现苹果植株并没有表现出对相应病菌的抗性。PR蛋白基因不仅具有一定的抗病功能,同时对植物生长、抵抗逆境胁迫、抗植株衰老及生长调节剂诱导等方面同样发挥着至关重要作用。
本试验利用生物信息学方法对BpPR1的基因结构、启动子顺式作用元件、理化性质及同源性等进行分析比较。结果表明,BpPR1的cDNA全长为816 bp,CDS为531 bp,编码176个氨基酸,理论等电点为8.93,为碱性蛋白质,理论上具有较高的抗菌性。对其同源性进行比对,BpPRI与葡萄、枣亲缘关系较近。BpPR1基因结构中含有3个内含子和2个外显子,与拟南芥、毛果杨等PR1基因结构有所不同,但其编码区大小差别不大。PR蛋白是受多基因调控的蛋白质,不同植物的表达量存在相对差异。对启动子顺式反应原件进行分析得到,BpPRI中含有茉莉酸甲酯响应的顺式作用元件、光響应顺式作用元件、低温响应的顺式反应原件、脱落酸响应的顺式作用原件、MYBHvl结合位点、生长素响应的顺式作用元件、水杨酸响应的顺式作用元件、细胞分裂素的顺式作用元件等,说明BpPRI参与多种植物生长调节剂的应答,该蛋白如何通过生长调节剂响应调控白桦抗病性还有待进一步研究。
参考文献:
[1]李萌,哈努拉·塔斯肯,陈卓,等白桦4CL基因启动子克隆及表达分析[J].中国农学通报,2017,33(5):29-34
[2]王家啟,张曦,李莉,白桦HD-Zip基因家族生物信息学及应答盐胁迫分析[J].植物研究,2018,38(6):931- 938.
[3]申婷婷,姜静,刘桂丰,等白桦BpCesA基因的生物信息学及表达分析[J].植物研究,2016,36(6):909-916.
[4] YANG C P Research progress on genetic improvement of Betulaplatyphylla SukEJ] Front Agr Sci Eng, 2017 .4(4): 391-401.
[5]佟志鹏,安梦楠,丁铖松,等.植物病程相关蛋白PR-NP24研究进展[J].分子植物育种,2019( 11):3542-3548.
[6] FARRAKH S .WANG M N . CHEN X M. PaLhogenesis-related pro-tein genes involved in race-specific all-stage resistance and non-race specific high-temperature adult-plant resistance to Pu.cciniastriiformis f.sp.tritici in wh e at[ J ] . Journal of integrativ e agriculture ,2018.17(11) : 2478-2491.
[7] ZHANC G. LI Y M, ZHANC Y. et al. Cloning and characterizationof a pathogenesis-related protein gene TaPRIO from wheat inducedby stripe rust pathogen [Jl. Agricuhural sciences in China. 2010.9( 4) : 549-556.
[8]张玉,杨爱国,冯全福,等,植物病程相关蛋白及其在烟草中的研究进展 [J].生物技术通报 , 2012( 5 ) : 21-23.
[9] L1 R H . CHEN S. LIU G F. eL al. Characterization and idenLifcationof a woody lesion mimic mutant Imd , showing defence response andresistance to Alternaria alcernate in birch [Jl. Science citation in-dex.2017.9: 1-12.
[10] SALOJARVI J. SMOLANDER O P. NIEMINEN K. el al. Cenomesequencing and population genomic analyses provide insights inLo the adaptive landscape of silver birch [J] . Nature genetics. 2017 ,49(6) :904-912.
[11] HU B. JIN J. GUO A Y. GSDS 2.0 : An upgraded gene ieature vi-sualizaLion serverLJl . Bioinformattics.2015 .31 ( 8) : 1296-1297.
[12] LE H G. HEITZ T. MESTRE P. et al. Characterization of Vitis vi-nifera NPRI homologs involved in che regulation of pathogenesis-related gene expression[J].BMC plant biology, 2009,9:54-64.
[13] SABATER J A B,ALMACRO L,BELCHI N S,et al Induction ofsesquiterpenes, phytoesterols and extracellular pathogenesis-relat-ed proteins in elicited cell cultures of Capsicum annuum[Jl. PlantPhysiol. 2010. 167 (15): 1273-1281.
[14]楊瑞瑞,易小娅,曾幼玲.PRlO的结构、信号转导以及功能的研究进展[J].中国农学通报,2014.30(4):251-258.
[15]王勇刚,曾富华,吴志华,等,植物诱导抗病与病程相关蛋白[J]湖南农业大学学报(自然科学版),2002.28(2):177-182.
[16]侯丽霞,高超,车永梅,等葡萄病程相关蛋白l基因的克隆和表达分析[J].植物生理学报,2012. 48(1):57-62.
[17]马立功,张匀华,孟庆林,等.向日葵病程相关蛋白HaPRl基因的克隆与功能[J].作物学报,2015 .41(12):1819-1827.
[18]张凯敏,王玉成,杨桂燕,等,柽柳TpPRI基因的克隆与表达分析[J].南京林业大学学报(自然科学版),2013. 37(2):45-49
[19] 11 Z T,DHEKNEY S A. CRAY D J PR-I gene family of grape-vine: Auniquely duplicated PR-I gene from a Vitis interspecifichybrid confers high level resistance to bacterial disease in trans-genic LobaccoLJl. Plam Cell Rep. 2011, 30: l-ll.
[20] BONASERA J M, KIM J F,BEER S V.PR genes of apple: Identi-fication and expression in response to elicitors and inoculation with Erwinia amvlovora[J]. BMC Plant Biol, 2006,6:23-34.
基金项目:国家自然科学基金项目(31800558)
作者简介:安玉婷(1991-),女,黑龙江安达人,在读硕士研究生,研究方向为植物学,(电话)18904534279(电子信箱)swxanyuting@126.com;通信作者,李然红,(电话)18504539007(电子信箱)swxlrh@126.com。
猜你喜欢
知音(月末版)(2020年5期)2020-07-04
中国校外教育(下旬)(2016年11期)2016-12-27
中国教育信息化·基础教育(2016年10期)2016-12-20
今传媒(2016年11期)2016-12-19
华人时刊(2016年16期)2016-04-05
