
水稻黄叶突变体yl6(t)的遗传分析和基因定位探究
2020-08-19 13:00袁铠
南方农业·中旬 2020年6期
关键词:水稻
袁铠
摘 要 在粳稻品种日本晴群体中发现了1个黄叶自然突变体yl6(t),因此,对其进行调查研究。农艺性状调查发现,与野生型相比,yl6(t)的结实率明显较低;叶绿素含量测定发现,突变体yl6(t)苗期后期叶素绿含量极显著降低,全生育期为黄叶;超微结构观察发现,叶绿体形态改变,类囊体片层结构减少;遗传分析表明,yl6(t)黄叶表型为单隐性核基因控制,利用SSR标记将目的基因初步定位在水稻第6号染色体约56 kb区段内,该区段未有相关基因报道,推测是1个新的控制水稻叶色的基因。
关键词 水稻;黄叶突变体;遗传分析;基因定位
水稻(Oryza sativa L.)是世界上三大粮食作物之一,中国绝大多数人口以水稻为主食。随着人口的增加和环境的恶化,提高水稻的产量显得十分重要。水稻产量的95%来自叶片叶绿体的光合作用,叶片是植物光合作用的主要器官。因此,研究水稻的叶色有重要的意义。
本研究对从日本晴群体中发现的自然突变的黄叶突变体进行了农艺性状考察,对其进行遗传分析和基因初步定位,为进一步精细定位和克隆该基因奠定基础。
1 材料与方法
1.1 供试材料
黄叶突变体由粳稻日本晴经自然突变而来,暂命名为yl6(t),该突变体经多年种植发现突变性状能稳定遗传。其他水稻材料包括正常叶色粳稻品种日本晴和籼稻品种Kasalath。亲本材料种植于扬州大学水稻试验田,遗传分析和定位群体种植于扬州大学农学院酒甸实验基地,均按照常规水肥管理。
1.2 叶绿素含量测定
于苗期、分蘖期及成熟期分别测定突变体yl6(t)和野生型日本晴的叶绿素含量。各取3株的倒2叶设定生物学重复,称取伸展完全的鲜叶(去叶脉)0.2 g。参照Lichtenthaler等的方法[1],用分光光度计分别测定664 nm、646 nm、470 nm波长处的吸光值D,进行叶绿素a、叶绿素b与类胡萝卜素含量测定。
1.3 叶绿体超微结构观察
在水稻苗期、分蘖期分别取突变体yl6(t)与野生型日本晴的叶片,在1~3 min取样,用刀片切成2 mm×2 mm的矩形小片,经过生物公司处理制成超薄切片,观察突变体和野生型叶肉细胞的叶绿体形态并拍照[2]。
1.4 主要农艺性状调查与分析
成熟期分别随机选取两亲本各6株,统计其剑叶长、剑叶宽、有效穗数、饱粒数、总粒数、结实率、主穗长、一次枝梗数、二次枝梗数、千粒重、株高,计算平均值,进行t测验。
1.5 遗传分析群体和基因定位群体构建
以正常日本晴为父本与突变体yl6(t)进行杂交得到F1,自交获得F2,以F2为遗传分析群体。以Kasalath为父本与突变体yl6(t)杂交F1,自交获得F2,从F2中分离黄叶表型的单株作为基因定位群体。
1.6 突变体yl6(t)的基因定位
采用BSA法,建成黄叶突变池和正常池。利用SSR标记对两个亲本突变体yl6(t)与Kasalath进行多态性分析,利用多态性良好的标记分析亲本及池间的多态性,找出与目的基因可能连锁的标记,利用隐性黄叶群体进行验证,确定目标基因所在染色体。
根据目标基因所在的位置,设计STS标记并挑选出多态性良好的标记,利用突变体yl6(t)与Kasalath的F2群体中的1 430株黄叶突变单株进行精细定位,DNA的提取采用CTAB法。
PCR反应过程:94 ℃,5 min预变性;95 ℃,40 s变性;51~56 ℃,40 s退火;72 ℃,40 s延伸,35个循环(潮霉素30个循环);72 ℃,10 min;16 ℃保存。
2 结果与分析
2.1 突变体yl6(t)的表型特征
通过观察种植在扬州的野生型日本晴和突变体yl6(t)可以明显发现,与野生型日本晴相比,突變体yl6(t)在苗期后期表现为淡黄叶,生长期间不转绿,成熟期时植株叶色已明显黄于野生型。突变体株高较矮,分蘖较少。
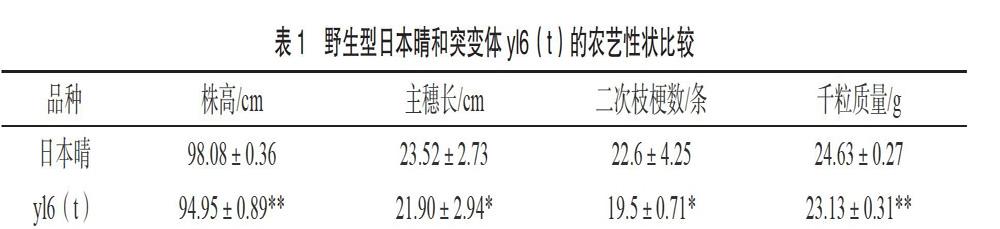
突变体yl6(t)与野生型日本晴相比,农艺性状也有所改变,具体见表1。两者二次枝梗数、主穗长有显著差异,株高、千粒质量达到了极显著差异,其他农艺性状差异不显著。
2.2 突变体yl6(t)的叶绿素含量测定
用紫外线分光光度计分别测量了突变体yl6(t)和野生型日本晴叶片各时期的叶绿素含量。发现突变体yl6(t)苗期和分蘖期叶绿素含量均低于野生型日本晴。随着叶片的生长,突变体叶绿素a、叶绿素b和总叶绿素含量都极显著下降,而胡萝卜素含量显著下降,这说明突变体yl6(t)叶片变黄有可能是叶绿素含量减少导致的。
2.3 突变体yl6(t)的叶绿体超微结构分析
透射电镜观察发现:,与野生型日本晴相比,突变体yl6(t)的叶绿体形状不规则,内部含有嗜饿性板层小体,类囊体数量减少,说明叶绿体发育不正常。这些说明,突变体yl6(t)与野生型日本晴相比,不仅叶绿素含量有差异,叶绿体的发育情况也受到了影响,这些可能与叶色变异有关。
2.4 突变体yl6(t)的遗传分析与基因定位
以突变体yl6(t)为母本,与正常叶色日本晴杂交,F1代自交获得F2,在F2代中,苗期后期发生叶色的性状分离。随机调查500株,考察突变株和正常株数量,其中突变株数量为109株,正常株数量为391株,X2=2.56<3.84=X20.05,该结果符合孟德尔单基因分离比3∶1,证明黄叶突变体yl6(t)受一对隐性核基因控制。
利用480个SSR标记对两个亲本进行多态性分析,利用多态性较好的标记检测混池的多态性,发现标记RM5851有明显的多态性,位于6号染色体上,接着利用RM5851对隐性单株群体进行群体验证,跑胶发现突变体的带型与大部分隐性单株的带型相同,经过计算发现这个标记与目的基因的交换率约为10%,初步确定该黄叶基因位于第6号染色体上。为了进一步定位该黄叶基因,在标记RM5851附近发展了十几个标记,最后挑选出多态性良好的IDL标记:标记ID1279、ID1413、ID1505、ID1732。因为还有一批黄叶隐性单株群体,为了尝试进一步缩小定位区间,于是接着利用480株黄叶突变单株进行分析,将该基因定位于标记ID1279与ID1732之间,接着继续利用950株单株进行分析,结果发现,该基因与标记ID1437共分离,且在标记ID1432与标记ID1505分别有12个和15个交换单株,标记ID1413有18个交换单株,标记ID1610有37株交换单株,标记ID1499有13个交换单株,通过交换株个数的变化情况,表明该基因存在于标记ID1432与标记ID1499之间,物理距离约为56 kb。
3 结论与讨论
本研究中所采用的材料为自然突变而来的黄叶突变体,突变方式较为安全,经过测定发现:突变体yl6(t)的农艺性状发生了变化,叶绿素含量与野生型相比明显降低,叶绿体产生了变异、结构发生了改变。因此该黄叶突变体叶色发生变异可能是由于叶绿素含量变化和叶绿体发育异常共同引起的。
本研究将yl6(t)基因定位到水稻6号染色体56 kb区段内,该区段中未见有关黄叶基因的有关报道,因此推测yl6(t)是一个新的黄叶基因,对该基因进一步研究将有利于阐明水稻叶色变异的分子机制。
参考文献:
[1] Lichtenthaler HK.Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes[J].Methods in enzymology,1987,148C:350-382.
[2] 邱义兰,李红,彭克勤,等.水稻“斑马叶”突变体B411叶绿体超微结构的观察[J].作物学报,2010,36(1):184-190.
(责任编辑:赵中正)
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
格言·校园版(2021年30期)2021-07-06
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2020年1期)2020-02-11
农民致富之友(2019年30期)2019-10-21
农民致富之友(2019年23期)2019-08-16
农民致富之友(2019年14期)2019-05-24
新农村(2018年29期)2018-04-02
农产品市场周刊(2017年35期)2017-10-25
