
辣椒炭疽zf-C2H2基因家族的生物信息学分析
2020-08-18 12:16项孙寰
农业技术与装备 2020年7期
项孙寰
(湖南农业大学植物保护学院,湖南 长沙 410128)
辣椒是许多国家重要的经济作物,具有重要的药用与食用价值。辣椒炭疽病严重危害作物生长,对作物品质、产量有重要影响。炭疽菌侵染辣椒寄主的致病机制涉及多种转录因子的复杂基因调控过程。锌指类蛋白转录因子对基因调控起重要作用。自1831年Corda提出炭疽菌属(Colletotrichum Corda)后,越来越多的植保工作者对炭疽菌进行了研究,表明其地理分布和寄主范围均非常广泛[1]。炭疽菌菌体较粗大,在有氧、温度适宜(25℃~30℃)条件下可形成椭圆形芽孢。德国医师兼科学家罗伯·柯霍(Heinrich Hermann Robert Koch)首先于1870年从人体中分离出炭疽菌,从此生物学家开始对炭疽菌进行深入研究。炭疽菌(anthracis)通常以内孢子的形态出现在土壤里,是一种棒状的革兰氏阳性菌,可在草莓、苹果、辣椒、梨、枣、柑橘和虎耳草等多种植物中引发疾病,并对这些植物造成严重危害[2]。炭疽菌主要侵害果实和叶片,导致果实整体腐烂,果皮变黑色[3]。植物炭疽病全年均可发生,主要发生在植株较大、通风不良或管理失当时,但以高温多湿时较为严重。林春花等[4]研究表明,炭疽菌主要有胶孢炭疽菌复合群(C.gloeosporioides species complex)和尖孢炭疽菌复合群(C.acutatun species complex),其中C.siamense胶孢炭疽菌复合群为主要类群。研究表明,炭疽菌会在侵染寄主植物的过程中合成大量的效应蛋白,抑制寄主细胞的PTI(天然免疫)反应,利于病原菌的侵染,说明效应蛋白在炭疽菌的致病过程中起着关键作用[5]。据报道,胶孢炭疽菌和尖孢炭疽菌均可以引起橡胶树炭疽病,但胶孢炭疽菌的致病性远强于尖孢炭疽菌的致病性[6]。
炭疽菌中存在许多转录因子,对其转录过程具有重要作用。转录因子是一类蛋白质分子,能与基因的5’端特定序列专一性识别并结合,从而保证基因表达的顺利进行。生物学者多以拟南芥进行转录因子研究,Riechmann等人在拟南芥中鉴定出 1 500 个转录因子,包括 NAC、b-ZIP、Myb/c、WRKY 和AP2/ERF家族,其中对AP2/ERF家族的研究最为完善[7],AP2/ERF家族主要特征是具有高度保守的约60~70个氨基酸组成的AP2结构域[8]。锌指类蛋白是一种具有手指状结构域的转录因子,由Zn2+与氨基酸整合形成,在基因调控方面具有重要的意义。1983年,第一个具有基因转录调控作用的锌指蛋白TF3A在爪蟾卵母细胞中被发现,此后锌指类基因调控因子蛋白被广泛研究[8]。根据锌指蛋白中组氨酸(His)与半胱氨酸(Cys)残基个数与位置的不同,Berg等[9]将锌指蛋白分为C2H2、C2HC、C6、C8、C2HC5、C3HC4、C4HC3、C2C2 和 CCCH等9类。
本研究着眼于与辣椒炭疽病致病机制相关的重要转录因子zf-C2H2,利用生物信息学方法,获得zf-C2H2的基因家族序列,对该家族成员进行序列比对,分析其理化性质、蛋白质二级及三级结构、系统进化关系、进行亚细胞定位等,为进一步解析zf-C2H2对辣椒炭疽的致病机理提供一定科学依据。
1 材料与方法
1.1 辣椒炭疽zf-C2H2基因家族鉴定及蛋白序列分析
为了鉴定zf-C2H2基因家族成员,首先从NCBI中下载辣椒炭疽全基因组序列,从网站(http://www.transcriptionfavtor.org/Download/pfam_18.dbds.v2.03.txt)中下载Pfam结构域模型。以pfamscan扫描辣椒炭疽全基因组序列,将包含上述pfam domain的辣椒炭疽序列提取出来。以hmmscan(https://www.ebi.ac.uk/Tools/hmmer/search/hmmscan)进行预测鉴定,确保序列均含有zf-C2H2保守结构域,对这些序列归类,最终得到19条包含zf-C2H2的基因家族序列。再使用ExPASy-ProtParam(https://web.expasy.org/protparam/)在线工具计算相应蛋白序列的分子量、等电点和氨基酸数。使用TargetP(http://www.cbs.dtu.dk/services/TargetP/)对获得的蛋白序列进行亚细胞定位分析。1.2 zf-C2H2基因家族系统进化分析
从NCBI中下载链霉菌zf-C2H2蛋白序列,利用MEGA X软件中的ClustalW功能对获得的蛋白质序列与辣椒炭疽zf-C2H2蛋白序列进行多序列比对,将得到的结果输入MEGA X软件中构建进化树,以此对基因的亲缘关系进行探究。
1.3 zf-C2H2蛋白二级、三级结构分析
辣椒炭疽zf-C2H2蛋白利用 SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)进行二级结构预测对zf-C2H2蛋白。利用SWISS-MODLE(https://swissmodel.expasy.org/interactive)进行三级结构预测。基于预测结果对蛋白质二级、三级结构进行分析。
1.4 zf-C2H2基因家族保守基序分析
一个基因家族中共有的motif很有可能是行使相应功能的关键序列或是基因家族中必不可少的序列。利用在线工具MEME(http://meme-suite.org/tools/meme)对 19条辣椒炭疽zf-C2H2蛋白的保守基序进行预测与分析。软件参数motif数目设置为8,其余参数使用默认值。
2 结果与分析
2.1 辣椒炭疽zf-C2H2基因家族成员理化性质及亚细胞定位分析
本研究从NCBI上下载辣椒炭疽基因蛋白序列,用pfamscan扫描Pfam-A,对所得80条序列MegAlign软件预测同源性,经hmmscan进行结构域鉴定,最后筛选出19条均含有zf-C2H2蛋白二聚区结构域的辣椒炭疽序列。将鉴定所得基因家族序列按照其基因ID排序从Cg1~Cg19命名。理化性质分析结果显示(表1),zf-C2H2蛋白的氨基酸数目在386~985aa,Cg5和Cg11的氨基酸数目最多;分子量最小的蛋白为Cg2和Cg10(42002.8KD),最大的蛋白为 Cg5(109661.9KD);蛋白质等电点在 6.05~10.04,其中仅 Cg1、Cg5、Cg11、Cg13 的等电点小于7(酸性蛋白),而其他15个家族成员均为碱性蛋白质。通过亚细胞定位分析可知,所有zf-C2H2蛋白都定位于细胞核中,推测zf-C2H2家族蛋白主要参与核基因组转录调控。
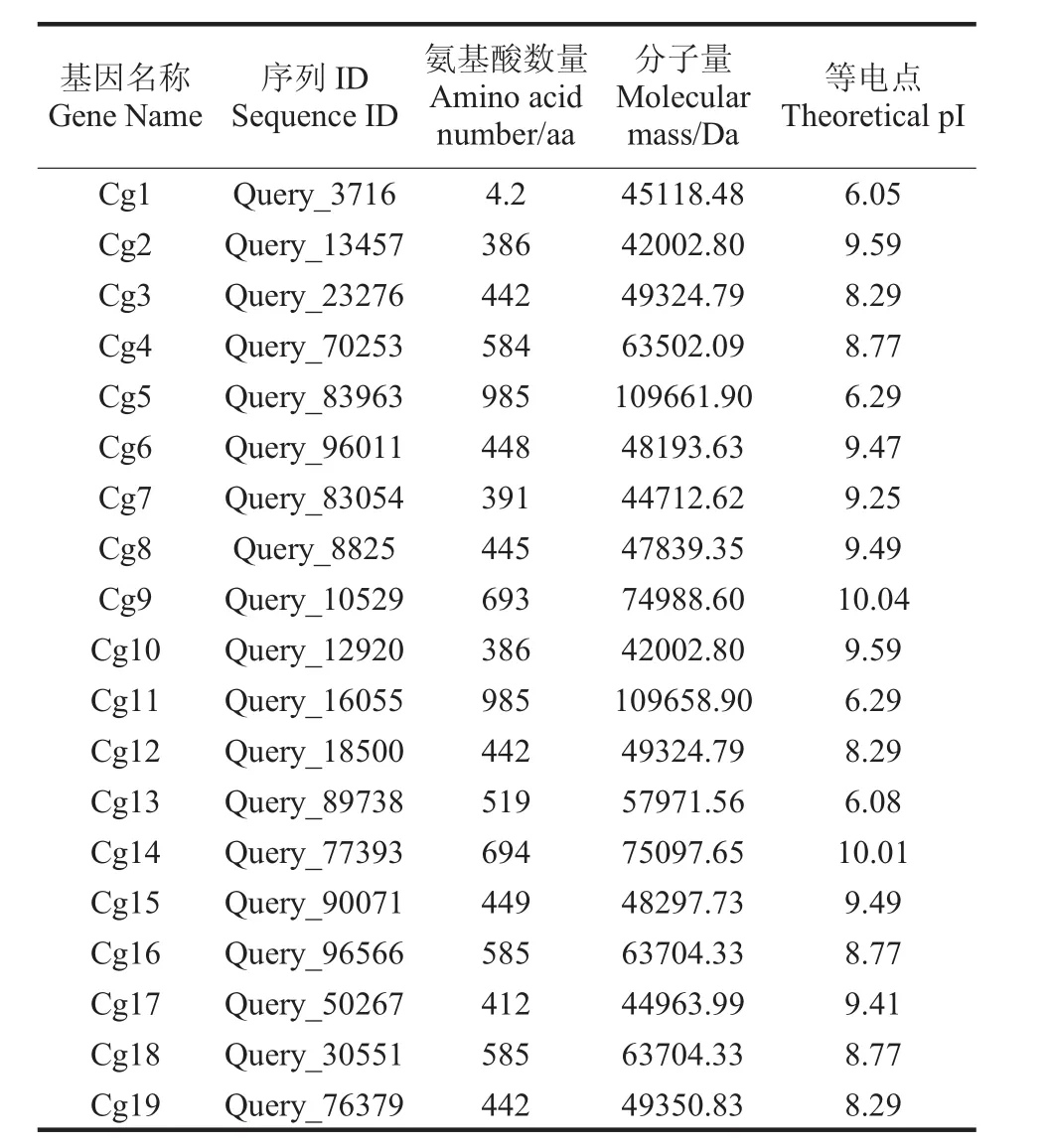
表1 辣椒炭疽zf-C2H2基因家族理化性质概况Tab.1 Physicochemical properties of zf-C2H2 gene family of Colletotrichum capsici
2.2 辣椒炭疽菌与链霉菌zf-C2H2基因家族进化分析
将辣椒炭疽zf-C2H2基因家族成员与链霉菌zf-C2H2基因家族进行多序列比对,之后利用MEGA X软件构建进化树(图1)。结果表明,辣椒炭疽zf-C2H2蛋白与链霉菌zf-C2H2蛋白家族位于不同分支,推测二者同源性较低;辣椒炭疽zf-C2H2蛋白分为7个亚族,第4与第7亚族的蛋白数量为2,其余亚族均包含3个蛋白。进化树显示,第7亚族(包含Cg5与Cg11)与链霉菌亲缘关系接近。
2.3 zf-C2H2基因家族蛋白结构分析
蛋白质二级结构是多肽链沿着主链骨架的空间布局、规则性循环式排列,或某一段肽链的局部空间结构,与氨基酸残基侧链的构象无关。蛋白质三级结构即在二级结构基础上进一步盘曲折叠而形成的三维空间结构,是肽链中所有氨基酸残基的相对空间布局。对辣椒炭疽zf-C2H2蛋白的二级结构预测分析表明(表2、图2),二级结构主要包括4种,即α-螺旋、β-折叠、延伸链与无规卷曲。其中无规卷曲所占比例最高,占57.46%~81.14%;β-折叠所占比例最少,仅占0.97%~4.75%。根据三级结构预测分析(见图3),可大致分为4组,一组为Cg1、Cg3、Cg7、Cg12、Cg13、Cg19;一组为 Cg2、Cg9、Cg10;一组为 Cg5、Cg6、Cg8、Cg11、Cg15;一组为 Cg4、Cg14、Cg16、Cg17、Cg18。每一组内的三级结构具有较高相似性,而组与组之间存在较大差异。

表2 辣椒炭疽zf-C2H2蛋白二级结构主要组件比例%Tab.2 Proportion of major components of zf-C2H2 protein secondary structure of Capsicum anthracnose
2.4 zf-C2H2基因家族保守基序分析
利用MEME在线工具对辣椒炭疽zf-C2H2基因家族蛋白进行保守基序查找,共搜索到8个motif(图4)。各motif的长度、氨基酸组成以及在家族成员中出现的次数等信息见表3,其中motif1在19条序列中均存在,而motif6仅存在于5个家族成员。其中 motif1、motif2、motif3、motif7 的保守性最高,且motif1、motif2、motif3这3种原件普遍存在于19个成员中。推测motif1、motif2、motif3、motif7 是辣椒炭疽 zf-C2H2 基因编码蛋白的重要组件。在同一亚族的zf-C2H2蛋白保守基序组成相似,验证了所鉴定出的辣椒炭疽zf-C2H2基因家族序列的可靠性。
3 讨论
锌指蛋白(如zf-C2H2)转录因子在自然界中分布广泛,且在基因调控网络中扮演重要角色。C2H2型锌指蛋白是最常见的锌指蛋白之一,对真核生物的生长发育及逆境胁迫的耐受能力都有着重要作用[10]。杨明磊等人[11]在分析烟草C2H2锌指蛋白转录因子家族成员时指出,C2H2锌指蛋白转录因子家族编码的转录因子具有多种生物学功能,其不仅与DNA、RNA相互作用,还与蛋白质相互作用。目前国内外对植物中尤其是拟南芥中的zf-C2H2蛋白分析较多,而对细菌内存在的zf-C2H2蛋白研究较少。炭疽菌等细菌通过寄生作物而导致作物品质、产量大幅下降。辣椒具有重要的药用及食用价值,研究炭疽菌对其侵染寄生机制有助于辣椒种质创新与育种改良。本研究采用生物信息学方法对辣椒炭疽zf-C2H2基因家族进行了鉴定及分析,旨在为炭疽菌中锌指蛋白参与的基因调控与寄生机制提供一定的科学依据。
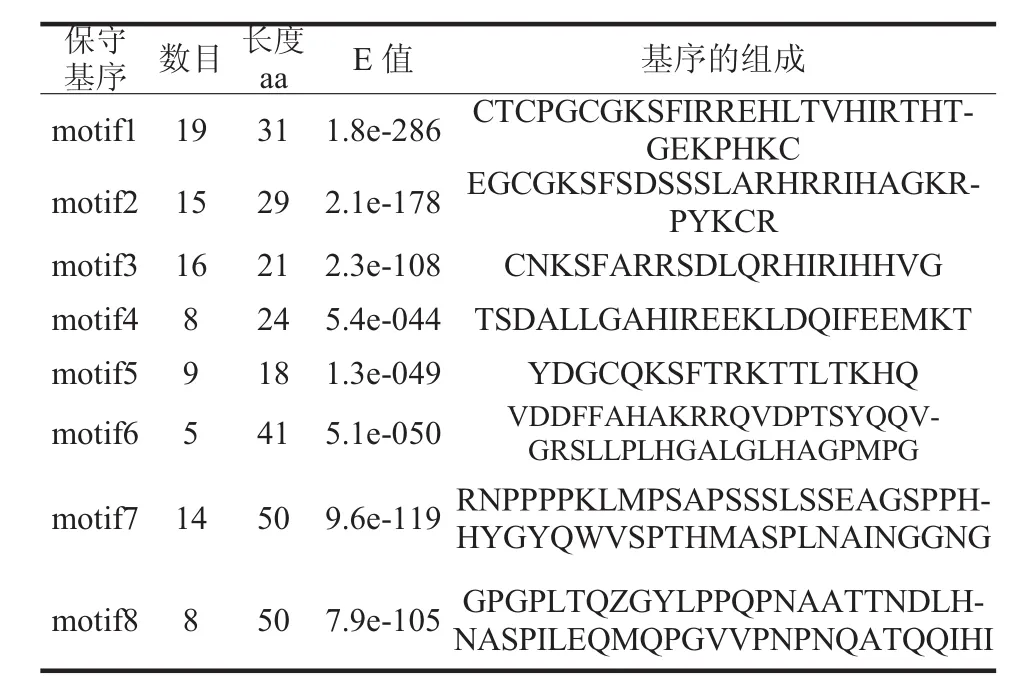
表3 基序的氨基酸组成Tab.3 Amino acid composition of motif
本研究用辣椒炭疽全基因组序列比对pfam结构域,去除掉不含有zf-C2H2结构域的基因,共筛选出19条辣椒炭疽zf-C2H2基因家族序列。通过对家族成员蛋白理化性质进行分析,发现zf-C2H2蛋白多为碱性蛋白,以碱性氨基酸为主,其中氨基酸数量与分子量差异不明显,表明基因家族成员之间具有相似的理化性质,符合基因家族定义的逻辑认知。通过亚细胞定位分析发现19条序列均位于细胞核,表明zf-C2H2蛋白主要在细胞核中起作用,参与遗传信息表达与调控。通过对辣椒炭疽zf-C2H2蛋白二级结构和三级结构分析,其中蛋白质二级结构无规卷曲占比大,其余结构占比小。按各结构类型所占比例大小排序为无规卷曲>α-螺旋>延伸链>β-折叠,表明无规卷曲结构在zf-C2H2蛋白二级结构中起着重要作用,其余结构则起到一定的修饰作用,同时,高含量的无规卷曲结构为解析转录因子结合位点及模式增加了难度。分析三级结构将序列大致分为4组,且同源性高的大多分在同一组,此现象验证了系统进化分析。由预测生成的三级结构与二级结构一致。近年来,多基因系统进化分析在炭疽菌分类研究中得到广泛应用[12-13],本文通过系统进化分析,发现辣椒炭疽zf-C2H2基因家族分为7个亚族,每个亚族的组成与数量都存在差异,推测差异源于基因的复制与丢失,但各个亚族之间的差异不明显,说明各亚族进化水平相差不大。保守基序分析验证了序列的系统进化分析,并且分析发现motif1存在所有分析序列中,推测motif1作为结构域中的亚单元,在结构域发挥多种生物学作用中具有重要意义。
目前C2H2型锌指蛋白在大豆、拟南芥、矮牵牛和水稻中均有较为系统且详尽的研究,但在其他物种研究方面相对欠缺。随着生物信息学的发展以及相关技术的进步,对该分子的研究会更全面。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
方圆(2021年19期)2021-11-03
小哥白尼(神奇星球)(2021年6期)2021-07-28
小学生必读(中年级版)(2021年12期)2021-03-03
小学生必读(中年级版)(2021年11期)2021-02-22
实用肿瘤学杂志(2020年4期)2020-12-08
快乐作文(1.2年级)(2020年8期)2020-09-10
中外文摘(2020年9期)2020-06-01
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
