
白色念珠菌生物膜与Toll样受体通路的免疫关系
2020-08-13 09:11赵冬梅冯奇刚张海云
牡丹江医学院学报 2020年3期
赵冬梅,王 琨,冯奇刚,张海云
(1.牡丹江林业中心医院检验科,黑龙江 牡丹江 157011;2.南通市第六人民医院检验科,江苏 南通 226000)
白色念珠菌是引起血流感染最常见的致病菌之一,其致死率不断升高[1-2]。随着导管插管、人工植入装置的不断应用,机体在感染白色念珠菌以后菌株容易粘附在表面形成生物膜[3-5],对各种环境刺激,如过氧化物、PH改变的敏感性降低[6-8],耐药性将上升10~1000倍[9-10]。通常情况下,在人体感染白色念珠菌之后,表达于树突状细胞、巨噬细胞、中性粒细胞等固有免疫细胞的模式识别受体—Toll样受体(Toll-like receptors,TLRs),通过识别不同致病菌的相关分子模式(pathogen associatedmolecμLarpatterns,PAMPs)激活固有免疫细胞及特异性免疫反应,在感染早期发挥作用而将其杀死,但生物膜形成以后这些成分被包裹而发生免疫逃逸[11-12]。目前,临床上对于产膜白色念珠菌引起的感染缺乏有效的治疗措施。本文分析产膜白色念珠菌感染的患者机体Toll样受体通路的变化,探讨产膜白色念珠菌在患者体内的免疫逃逸机制,为临床治疗白色念珠菌所致感染性疾病提供理论依据。
1 材料与方法
1.1 研究对象收集2017年1月至2018年12月间从牡丹江林业中心医院住院血流感染患者(60例)标本中分离的白色念珠菌,根据白色念珠菌是否产生生物膜将相应病例分成产膜组和非产膜组,每组30例;另纳入30例健康人为健康对照组。产膜组中男18例,女12例,年龄20~80岁,平均年龄(55.25±6.41)岁。非产膜组中男16例,女14例;年龄25~81岁,平均年龄(58.37±7.63)岁。健康对照组中男17例,女13例,平均年龄(56.84±7.49)岁。纳入标准:(1)年龄18~80岁,资料完整者;(2)在血液培养中分离出白色念珠菌的患者(不包括健康对照组)(3)意识清晰者,生命体征平稳者。排除标准:(1)在血液培养中分离出其他病原菌者;(2)三天内服用抗真菌药物者;(3)合并心肝肺等脏器功能衰竭者。三组研究对象性别比例、年龄等资料无显著差异(P>0.05)。本研究经我院医学伦理委员会批准,患者及家属同意此次研究并签署相关知情同意书。
1.2 实验试剂与仪器结晶紫染料、RPMI 1640培养基、小牛血清(FBS)、RIzol(Invitrogen公司)、YPD液体培养基。RNA反转录试剂盒、ELISA试剂盒(上海齐一生物科技有限公司)、血清总蛋白提取试剂盒(普利莱基因技术有限公司)、基因引物由上海生工生物工程有限公司合成。荧光倒置显微镜、荧光定量PCR仪(Iqtm5,QLYMPUS)、酶标仪(680,Bio-Rad)、全自动血培养仪(美国BD公司)、二氧化碳培养箱、离心机等。
1.3 血液标本采集及白色念珠菌的分离分别采集产膜组、非产膜组与健康对照组外周静脉血5mL保存于EDTA抗凝管中,4℃冷藏备用。以白色念珠菌10231作为标准菌株(温州康泰生物)。当血流感染患者体温超过38℃时,无菌操作采集其静脉血10 mL注入血培养瓶中,于全自动血培养仪中培养。报阳后利用珠海迪尔公司全自动微生物分析仪(DL96Ⅱ)将微生物鉴定到“种”水平,分纯备用。
1.4 白色念珠菌生物膜体外模型的构建及分析将1.3中分离出的白色念珠菌接种于血培养皿,置于37℃二氧化碳培养箱培养24h,后用长棉签沾取单个菌落,混合在RPMI 1640培养基中,调整菌浓度为1.0×106/mL。取100μL菌悬液加到96孔细胞培养板中,置二氧化碳培养箱中培养72h,经PBS冲洗3遍,戊二醛固定,最后加入结晶紫染液,30 min后冲洗,酶标仪检测吸光度。评价标准:A570>0.25为产膜菌;A570<0.25为非产膜菌[1-2]。加入荧光染料FITC-conA后,在荧光显微镜下观察不同产膜能力的菌株形态。
1.5 RT-PCR检测各组外周血单个核细胞中TLR2、TLR4、Myd88基因的表达量采用Ficoll密度梯度离心法直接收集外周血单个核细胞,提取总RNA,qRT-PCR测定基因TLR2、TLR4、MyD88 mRNA表达水平(引物序列见表1)。以β-actin为内参基因,采用2-△△Ct相对定量计算方法分析目的基因表达量的变化。

表1 引物序列
1.6 Western Blot检测各组外周血单个核细胞中Myd88蛋白的含量按照1.5的方法收集单个核细胞,参照说明书配置总蛋白溶液,BCA法测定蛋白质浓度。经电泳、转膜、封闭和孵育后,用化学发光剂显影,获取图像,Image J 软件处理系统分析Myd88蛋白及β-actin管家基因的表达水平。
1.7 ELISA测定各组细胞因子的分泌情况将研究对象外周血液标本,离心取上层血浆,采用ELISA方法对各组血浆中细胞因子IL-1β、TNF、IFN-γ、IL-4的表达量进行测定(具体测定方法参照试剂盒说明)。
1.8 统计学方法采用SPSS 23.0统计学软件进行分析,计量数据资料采用“均数±标准差”表示,组间比较则采用单因素方差分析,进一步两两比较行LSD法。P<0.05为差异有统计学意义。
2 结果
2.1 荧光显微镜观察FITC-conA染色后的菌株形态荧光显微镜观察FITC-conA染色后的菌株,发现产膜菌呈菌丝状相互交错密集排列,而非产膜菌大部分以酵母样黏附于培养板底部,见图1。
2.2 各组外周血单个核细胞中TLR2mRNA、TLR4mRNA、Myd88mRNA相对表达量的比较患者感染白色念珠菌后,机体对产膜菌与非产膜菌的免疫应答能力不同。以健康对照组为基线并设值为1,产膜组外周血单个核细胞中TLR2mRNA、TLR4mRNA、Myd88mRNA的相对表达量相较健康对照组低,而非产膜组相较健康对照组高,差异具有统计学意义(P<0.05)。见图2。
2.3 各组外周血单个核细胞中Myd88蛋白含量的比较产膜组Myd88蛋白的含量较健康对照组显著降低(P=0.002),非产膜组较健康对照组显著升高(P=0.002),差异有统计学意义(P<0.05),见图3。

图3 各组外周血单个核细胞中MyD88蛋白的含量
2.4 各组血浆中细胞因子分泌情况的比较与健康对照组相比,产膜组患者血浆中IL-1β、TNF、IFN-γ浓度明显降低(P<0.05),非产膜组上述指标浓度明显升高,差异有统计学意义(P<0.05)。在IL-4因子浓度变化方面,产膜组与健康对照组相比明显升高(P<0.05),非产膜组与健康对照组相比差异无统计学意义(P>0.05)。见表2。
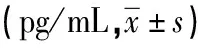
表2 ELISA测定各组细胞因子的浓度
3 讨论
有研究表明白色念珠菌生物膜是由大量菌丝相细胞、少量酵母相细胞以及由菌体分泌的基质共同组成的复杂结构[7-8]。因此,本实验将从血流感染患者中分离出的菌株分为产膜组和非产膜组,利用荧光显微镜观察其形态,发现产膜菌多呈菌丝状相互交错密集排列,而非产膜菌大部分以酵母样黏附于培养板底部,说明酵母相可能是白色念珠菌非生物膜状态的主要构成部分,菌丝相可能是白色念珠菌生物膜态的主要构成部分。菌丝态可维持成熟生物膜的结构的完整性和维持多层体系,具有抗吞噬作用,适应性和毒力更强,被认为更有利于逃逸宿主防御[12-13]。
在机体感染白色念珠菌时,模式识别受体TLR2和TLR4识别白色念珠菌表面的糖磷脂甘露聚糖,通过衔接蛋白Myd88,依靠Myd88依赖性途径和Myd88非依赖性途径来激活NF-κB和丝裂原蛋白激酶(mitogen-zctivatedprotein kinase,MAPK)级联反应通路,激活NF-κB信号传导通路,促进IFN-γ、IL-1、TNF-a等炎症因子的表达[14-15],诱导CD4+T细胞向Th1型细胞分化,介导细胞免疫应答[16-17]。Myd88是所有TLR家族通路中的必须分子,被称为“万能衔接蛋白”[18-19]。有研究显示,TLR4激活后,胞内衔接蛋白Myd88表达量增加;采用TLR4抗体阻断后,Myd88表达量减少[20],TLR2基因敲除抑制小鼠Myd88的高表达[21],Myd88基因敲除后,小鼠的TNF-α和IL-1β分泌水平降低[22]。所以TLR2mRNA、TLR4mRNA、Myd88mRNA的低表达,直接限制TLR/Myd88/NF-κB信号通路传导,减少促炎性细胞因子的释放。至于Myd88的缺失是否能反向降低TLRs表达,还需要有研究数据来证实。
本文实验数据显示,单核—巨噬系统在吞噬杀灭非产膜菌和产膜菌的过程中,TLRs mRNA、Myd88mRNA和细胞因子的表达量有显著差异,产膜组TLR2 mRNA、TLR4 mRNA、Myd88 mRNA/蛋白和细胞因子IL-1、IFN-γ及TNF-α的表达量明显低于健康对照组,而非产膜组表达量高于健康对照组,表示产膜菌TLR/Myd88/NF-κB信号通路受阻,促炎性细胞因子释放减少,Th1型免疫应答减弱;另外细胞因子IL-4的表达量也不同,产膜组IL-4的表达量明显高于健康对照组,启动辅助性T细胞由Th0分化为Th2,触发Th2型免疫应答,促进B淋巴细胞发育和介导体液免疫[23-24]。体液免疫对巨噬细胞、中性粒细胞的吞噬和杀伤病原体的能力有一定的抑制作用[23-24]。
综上所述,白色念珠菌生物膜可能通过下调TLR2的表达、干扰TLR4活化,抑制关键衔接蛋白Myd88的表达,使TLR/Myd88/NF-κB信号通路传递受限,无法诱导产生IFN-γ、TNF-α等促炎性细胞因子,使机体免疫应答朝着Th2型体液免疫反应发展,这可能是白色念珠菌逃逸机体免疫效应的一种方式,另一方面,也可能是机体调控白色念珠菌感染所致的炎症反应的重要机制。目前已经有研究表明,Ndt80是白色念珠菌形态转换中转录基因激活所必需,转录因子Tecl纯合缺失突变体使菌丝发育受阻[5,25-26]。下一步可以此为靶点研究药物针对基因的靶向治疗。随着分子免疫学和分子生物学技术的发展,基因工程抗体、分子疫苗、免疫佐剂、细胞因子和给予免疫激活等免疫技术的应用,治疗产膜白色念珠菌感染的难题一定会被攻克。
猜你喜欢
首都食品与医药(2022年11期)2022-06-06
现代临床医学(2022年2期)2022-04-19
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
生物技术通报(2021年2期)2021-04-26
临床检验杂志(2020年1期)2020-03-02
现代检验医学杂志(2016年1期)2016-11-12
中国科技信息(2015年2期)2015-11-16
郑州大学学报(医学版)(2015年1期)2015-02-27
现代检验医学杂志(2015年2期)2015-02-06
