
Eye location,cranial asymmetry,and swimming behavior of different variants of Solea senegalensis
2020-08-08 07:50JubinXingZhongheKeLipingLiuChenhongLiXiolingGongBolongBo
Aquaculture and Fisheries 2020年4期
Jubin Xing,Zhonghe Ke,Liping Liu,Chenhong Li,Xioling Gong,*,Bolong Bo,**
a Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources(Shanghai Ocean University),Ministry of Education,International Research Center for Marine Biosciences at Shanghai Ocean University,Ministry of Science and Technology,National Demonstration Center for Experimental Fisheries Science Education,Shanghai Ocean University,Shanghai,201306,China
b Fishery Technical Extension Station of Zhuji City,Zhuji,Zhejiang,311800,China
A R T I C L E I N F O
Keywords:
Flatfish
Eye migration
Left/right asymmetry
Senegalese sole
A B S T R A C T
Many flatfish species,such as Scophthalmus maximus,Paralichthys olivaceus,Solea senegalensis,are extensively cultured worldwide.In flatfishes,eye migration varies among different species and even within the same species.The genetic mechanism underlying eye migration is still unclear,and it is the key to improve metamorphosis in flatfish aquaculture and understand flatfish evolution.In this study,the high frequency of eye location variants in an artificial S.senegalensis population allowed us to investigate the eye preference during migration;completeness of the eye migration;and other related left-right asymmetric characteristics,such as asymmetrical skull remodeling and lying down on one side.The different frequencies of eye preference during migration,completeness of the eye migration,and settling side suggest that they are determined by different genotypes and imply independent evolutionary events during flatfish evolution.We found that the swimming time may be related to the completeness of eye migration,and further the migration of the eye,the more time the fish spent at the sea bottom.The left-right asymmetric degrees of not only the frontal bones and lateral ethmoids but also the jaw bone,including the premaxillary,maxillary,and dentary bones,depend on the completeness of eye migration.Finally,we proposed a hypothesis for the asymmetric origin of flatfish that provides reasonable explanations for the selective advantages of primitive flatfish species.
1.Introduction
Flatfishes(Pleuronectiformes)are generally deep-bodied and laterally compressed fishes,and they represent a diverse order of marine and estuarine fishes.Around 716 flatfish species can be found worldwide,and they comprise a significant proportion of the groundfish catch(Gibson,2005).Flatfishes are among the most popular and valuable fishes used for human consumption;therefore,they are attractive candidates for both stock enhancement and intensive cultivation.For example,the turbot(Scophthalmus maximus),olive flounder(Paralichthys olivaceus),and Senegalese sole(Solea senegalensis)have become important aquaculture species worldwide.
During the aquaculture of flatfishes,inherent variations in growth and settling behavior during metamorphosis may lead to cannibalism and necessitate increased labor due to grading(Gavlik,Albino,&Specker,2002;Gavlik &Specker,2004).To produce a larger and more uniformly sized cohort of juvenile flatfishes,methods such as salinity change and thyroid hormone manipulation have been used to synchronize the settling behavior,growth,and development of the summer flounder,Paralichthys dentatus,and spotted halibut,Verasper variegatus(Gavlik et al.,2002;Gavlik & Specker,2004;Tagawa &Aritaki,2005).Understanding the regulation mechanism of flatfish metamorphosis is the key for synchronizing these metamorphic events in artificial populations.
All flatfishes begin their life as pelagic,bilaterally symmetrical fishes.However,during larval development,flatfishes undergo a spectacular ontogenetic metamorphosis where one eye migrates from one side of the head to the other(Brewster,1987).After eye migration,flatfishes settle out of the water column and assume a benthic lifestyle.Recently,studies on the metamorphosis of flatfishes have reported information on the regulation of the signal pathway for eye migration,frontal bone deformation,dorsal fin elongation,and changes in body depth during metamorphosis and mechanism underlying the establishment of left/right asymmetric pigmentation after metamorphosis in flatfish(Bao et al.,2011;Campinho et al.,2018;Chen,Liu,Yao,Gao,&Bao,2017;Li,Zheng,Bao,& Berendzen,2013;Shao et al.,2017;Sun et al.,2015;Wei,Chen,Chen,& Bao,2017;Xu,Ke,Xia,He,& Bao,2016).However,the genetic basis for eye location and settling side in flatfishes is still unclear.In this study,the relatively high diversity of eye location and settling side in an artificial population of S.senegalensis helped us to understand the relationships among eye preference during migration,completeness of eye migration,and settling side.
2.Materials and methods
2.1.Fish maintenance
Artificially fertilized eggs were collected from farm-raised Senegalese soles(S.senegalensis)at Dahua Fisheries Farm(Laizhou,Shandong,China).Hatching and larval rearing occurred at 20°C in saltwater with 30 salinity.After 3 days after hatching(DAH),the larvae started to feed on the rotifer Brachionus plicatilis.The larvae at 9 DAH were transferred to our lab at Shanghai Ocean University(Shanghai,China)and maintained in saltwater at 20°C and 30 salinity.The larvae were fed with live brine shrimp(Artemia)nauplii until 32 DAH.All individuals(total 1396)at 24 DAH had completed metamorphosis and developed into juveniles with a bottom-dwelling lifestyle.
2.2.Classification of S.senegalensis individuals
Normal S.senegalensis is typically dextral(right-sided),and only the left eye shows migration(Dinis,Ribeiro,Soares,&Sarasquete,1999).In the present study,four types of S.senegalensis were recorded on the basis of which eye migrated,final position of the movable eye,and settling side:Type 1:left eye migrated,including Type 1-A(left eye moved just over the dorsal midline,and body laid down on the left side),Type 1-B(left eye stopped at the dorsal midline,and body laid down on the left side),Type 1-C(left eye stopped below the dorsal midline,and body laid down on the left side),and Type 1-C’(left eye stopped below the dorsal midline,and body laid down on the right side).Type 2:reversed eye variants with right eye migration,including Type 2-A(right eye migrated over the dorsal midline,very close to the target location,and body laid down on the right side)and Type 2-B(right eye stayed below the dorsal midline,and body laid down on the right side).Type 3:both eyes migrated and close to the dorsal midline,and body laid down on the left side.Type 4:both eyes experienced no migration,and body laid down on the left side.The frequencies of the various asymmetric phenotypes were determined by counting the individuals at the post-metamorphosis stage(28 DAH).
2.3.Swimming behavior
The post-metamorphic juveniles of S.senegalensis were maintained in glass tanks(40 cm×30 cm×25 cm)to observe their swimming behavior with a digital camera(SONY).The videos were edited using the software Avidemux 2.5.
2.4.Skeletal staining and photography
The post-metamorphic juveniles of S.senegalensis were fixed in 4% paraformaldehyde for at least 24 h.Alcian blue 8GX(A3157;Amresco)was used for cartilage staining,and alizarin red S(A5533;Amresco)for bone staining.The staining procedures were performed as described previously(Gavaia,Sarasquete,& Cancela,1999).The samples were observed under a dissecting microscope(SZX7;Olympus)and documented using the Image-Pro Plus 6.0 software(Media Cybernetics).The scale on each photograph was autogenerated by Image-Pro Plus 6.0 software on the basis of the magnification.
3.Results
3.1.Frequencies of the asymmetric phenotypes in the artificial S.senegalensis population
Representative individuals with different eye location and settling side in the artificial S.senegalensis population are shown in Fig.1.Thefrequencies of each phenotype in a total of 1396 juveniles are listed in Table 1.The frequency of settling side different from that of the normal juvenile was 8.6‰,including Type 2-A,Type2-B and Type 1-C’variants,whereas the frequency of specific eye migration different from that of the normal juvenile was 9.3‰,including Type 2 and Type 3 variants.The variety in the final location of the migrated eye can reflect the completeness of eye migration.The phenotype of incomplete eye migration was 66.63‰,after excluding the Type 2-A variants.
3.2.Cranial asymmetry in the different eye variants
In the S.senegalensis wild type,some pair of bones located in the front part of the head,such as the frontal bones and lateral ethmoid,premaxillary,maxillary,dentary,lachrymal,entopterygoid,and ectopterygoid bones,are distributed with a left-right asymmetry(Fig.2).In Type 2-A(reverse eye variants),these pairs of bones also present leftright asymmetry,but as a mirror image of the wild type(Fig.2).
The left-right asymmetric degree of some pairs of bones seems to be related to the migration distance of the eye.Some pairs of bones presented lesser left-right asymmetry than those of the wild type.In Type 1-A and Type 1-B variants with one eye migrated on or over the dorsal midline,bones such as the premaxillary,maxillary,and dentary presented a left-right asymmetry that was very similar to that of the wild type(Fig.2).In Types 1-C and 4,whose eyes do not move or reach the dorsal midline,these three pairs of bones present almost left-right symmetry(Fig.2).Although the pair of entopterygoids present leftright asymmetry in the wild type,it should be noted that the pair of bones seem to be no business with eye asymmetry,because they present a similar degree of left-right asymmetry in variants with different types of eye location(Types 1-A,1-B,1-C,and 4)(Fig.2).
3.3.Different variants with different activities in the artificial S.senegalensis population
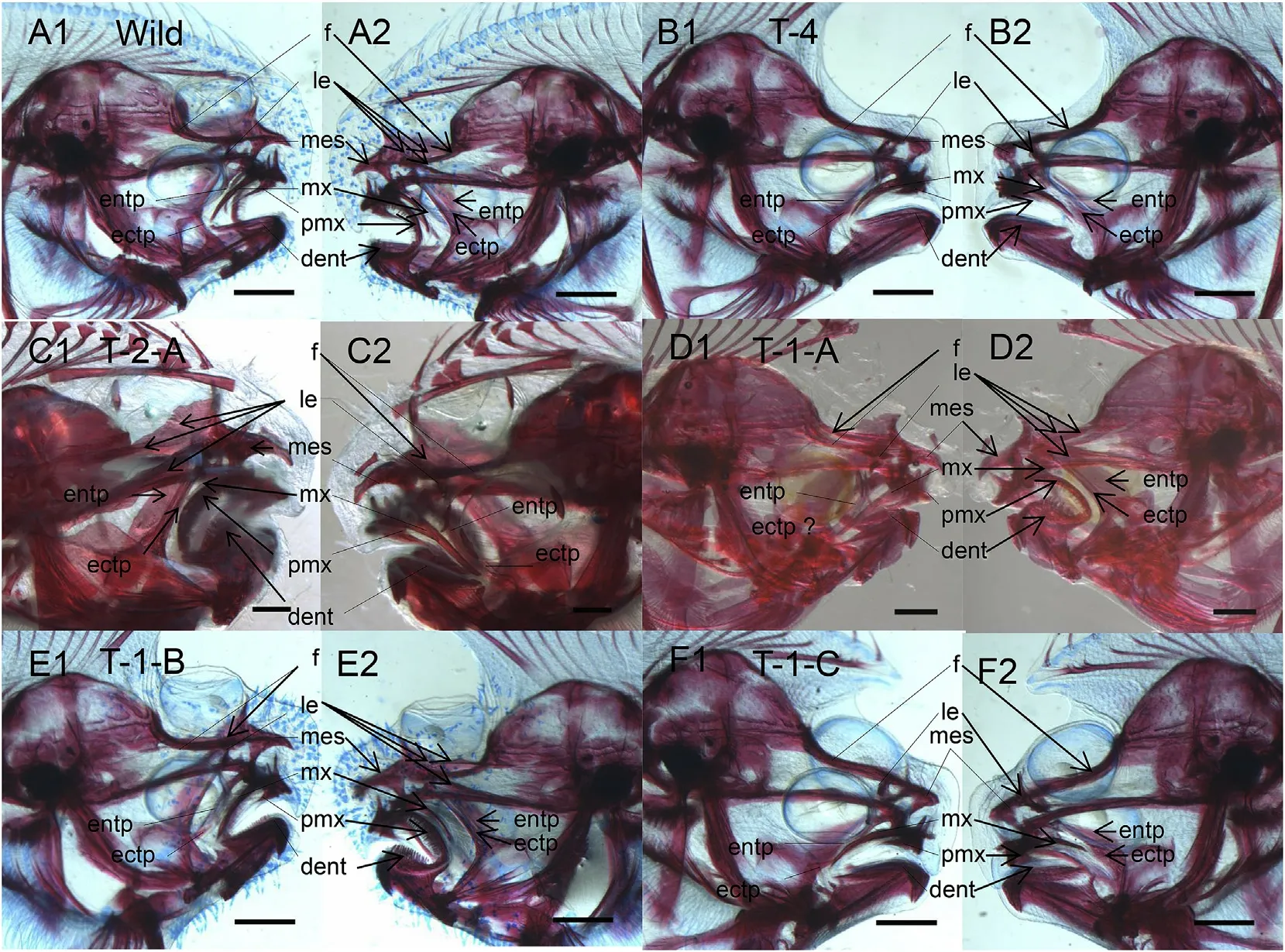
Fig.2.Skull staining for different variants with different eye locations.A,Wild type,(A1)Right view,(A2)Left view;B,Type 4,(B1)Right view,(B2)Left view;C,Type 2-A,(C1)Right view,(C2)Left view;D,Type 1-A,(D1)Right view,(D2)Left view;E,Type 1-B,(E1)Right view,(E2)Left view;F,Type 1-C,(F1)Right view,(F2)Left view.Bars,1.0 mm;f,frontal;le,lateral ethmoid;mes,mesethmoid;pmx,premaxillary;mx,maxillary;dent,dentary;entp,entopterygoid;ectp,ectopterygoid.Lines represent pairs of bones on the ocular side,and arrows represent pairs of bones on the blind side.Calcified bone is red,and cartilage is blue.
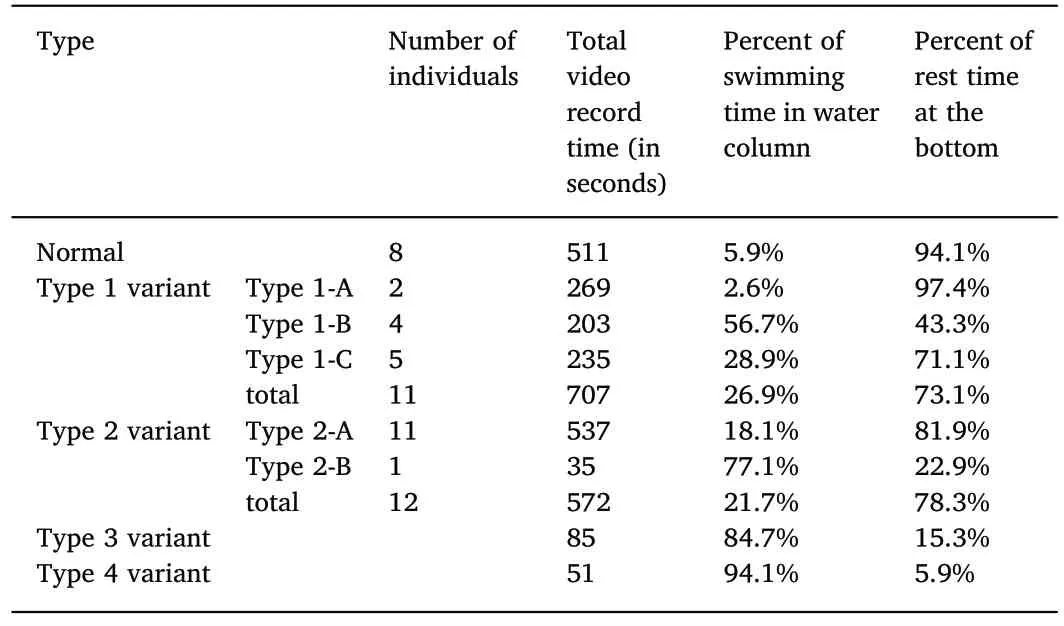
Table 2Different variants with different activities in the artificial S.senegalensis population.
The behaviors of the different variants seem to be related to the migration distance of the eye.The normal juveniles spent most of their time(94.1%)at the bottom on their eyeless body side(left side;Table 2).The Type 1 variants(Type 1-A,Type 1-B,and Type 1-C),with less left-eye migration,lay on the left side,swam upwards more often,and spent more time(26.9%)in water column(Table 2).The Type 2 variants(Type 2-A and Type 2-B),with less right-eye migration,lay on the right side and swam upwards more often(21.7%)in water column(Table 2).Type 1-A,with eye migrated over the dorsal midline,spent 97.4% of the time at the bottom,whereas Type 1-C,whose eye did not reach the dorsal midline,spent less time(71.1%)at the bottom(Table 2).The Type 3 variants,with both eyes close to the dorsal midline,spent most of the time(84.7%)in water column and only 15.3% of the time at the bottom.The Type 4 variants,with almost no eye migration,spent 94.1% of the time swimming(Table 2).
4.Discussion
In the artificial S.senegalensis population,the frequency of variants with incomplete eye migration was quite different from that of the variants with an eye preference for migration or different settling sides.Moreover,a Type 1-C’individual with mismatched eye migration and body settling directions showed that settling side and eye preference for migration are not determined by the same genotype,and even the frequency of variants with an eye preference for migration or different settling side was close.It is clear that these three phenotypes are determined by different genotypes and could be the result of independent evolutionary events during flatfish evolution.Schreiber(2006)suggested that tilting and settling behaviors are independent of asymmetric eye position on the basis of southern flounder variants produced by disrupting the thyroid hormone before or during metamorphosis.In our previous study(Bao et al.,2011),we proved that settling side is independent of eye migration in eye-symmetric S.senegalensis,Cynoglossus semilaevis,and P.olivaceus juveniles with a bottom-dwelling lifestyle;the specimens were produced artificially by inhibiting cell proliferation in the suborbital area on the blind side.
Swimming behavior can be affected by incomplete eye migration.The variants with incomplete eye migration,especially with eyes on both sides,might not live well in the wild because the swimming behavior of S.senegalensis was observed to be impaired by incomplete eye migration.Such specimens spent more time in water column than the normal individuals,suggesting that they would not adapt well to a benthic lifestyle.In contrast,the swimming behavior of reversed eye variants that showed complete eye migration(Type 2-A)was similar to that of normal individuals(that spend most of their time at the bottom),indicating these variants could adapt to a benthic lifestyle like the normal individuals.
Besides eye migration and settling behaviors,cranial asymmetry during flatfish metamorphosis is a striking asymmetric development in the vertebrates.Cranial asymmetry involves several bones,such as the frontal bone,lateral ethmoid,and pseudomesial bar(in some species)(Brewster,1987;Okada,Takagi,Seikai,Tanaka,& Tagawa,2001;Wagemans,Focant,& Vandewalle,1998).In the Japanese flounder,distortion of the frontal bone and left-right asymmetric enlargement of the lateral ethmoids during metamorphosis depend on eye migration(Bao et al.,2011;Sun et al.,2015).This study has also shown that,in S.senegalensis,left-right asymmetry of the frontal bone and lateral ethmoids is related to the migration distance of the eye and left-right asymmetric degree of the jaw,including the premaxillary,maxillary,and dentary bones,depends on eye migration.
The findings of this study suggest that the incompleteness of eye migration and settling side may play important roles in the evolution of flatfishes because the adaptive ability for a benthic ground lifestyle is mainly related to the completion of eye migration and flatfishes with the wrong settling side will not survive in the wild.Therefore,here we propose a hypothesis on the asymmetric origin of flatfish evolution.
It is unclear which living species is the closest to the ancestor of flatfishes;Perciformes fish species with compressiform bodies are supposed to be close to the ancestor of flatfishes(Dawkins,1986;Zimmer,2008).Upper binocular vision would be a very useful trait for a bottomdwelling species(Janvier,2008).A new species with asymmetric eyes,even if the eyes are still on both sides,could obtain upper binocular vision(Fig.3),like the fossil species Heteronectes and Amphistium with a possibly equal frequency of dextral and sinistral individuals(Friedman,2008).Obviously,the species with asymmetric eyes would have a more selective advantage than its ancestor with symmetric eyes.Regardless of which eye migrated,if the distance of eye migration is the same,the selective advantage should be the same.Therefore,the right or left eye could have equal opportunity to migrate in the same species.A species with one eye located at the dorsal midline,such as the spiny turbots(genus Psettodes;the most primitive living group that belongs to Pleuronectiformes)often lying on one side on sand and mud bottoms(Hussain,1990),acquires wider upper binocular vision and a more selective advantage than Heteronectes or Amphistium with eyes on both sides(Fig.3).Psettodes species spend most of their time lying at the bottom to maximize upper binocular vision and maintain front binocular vision at the horizontal level.In Psettodes erumei,the frequency of both eyes on the left or right side is similar in wild populations(Hussain,1990),suggesting that the preference and distance of eye migration are attributable to different genes.
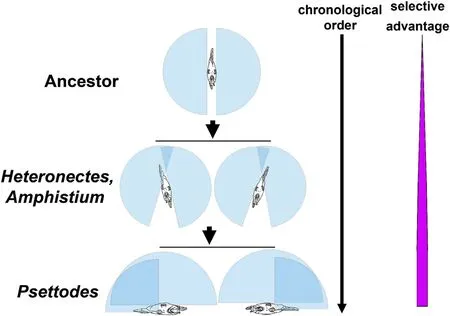
Fig.3.Scheme for the evolutionary origin of flatfish asymmetry.The ancestor of flatfishes from Perciformes is supposed to be compressiform with symmetric eyes.The upper vision field in each eye(light blue in the figure)is supposed to be 180°.Heteronectes and Amphistium species with asymmetric eyes that were still on both sides could have evolved from the ancestor because they obtained upper binocular vision(blue)and,accordingly,more selective advantage than their ancestor.The size of the pink area on the right side of the figure represents the selective advantage.Heteronectes and Amphistium could adjust the front binocular vision at the horizontal level via tilted swimming.The eye on the left or right side was upper lined with equal frequency in the same species,for example,Heteronectes,Amphistium,and Psettodes species.One eye located at the dorsal midline would help Psettodes species obtain a much wider binocular vision and,accordingly,selective advantage.
Ethical standards
The experiments comply with the current laws of China.
Declaration of competing interest
The authors declare that they have no conflict of interest.
Acknowledgements
This study was supported by the National Natural Science Foundation of China(31872546,31472262)and China-ASEAN Maritime Cooperation Fund through the project“China-ASEAN Center for Joint Research and Promotion of Marine Aquaculture Technology.”
 Aquaculture and Fisheries2020年4期
Aquaculture and Fisheries2020年4期
- Aquaculture and Fisheries的其它文章
- Genomes of major fishes in world fisheries and aquaculture:Status,application and perspective
- The economic contribution of fish and fish trade in Bangladesh
- Effect of chicken haemoglobin powder on growth,feed utilization,immunity and haematological index of largemouth bass(Micropterus salmoides)
- Effect of the number of freeze-thaw cycles number on the quality of Pacific white shrimp(Litopenaeus vannamei):An emphasis on moisture migration and microstructure by LF-NMR and SEM
- Hydrodynamic performance and structural response characteristics of the double-slotted vertical cambered V-Type otter board