
择时针刺足三里穴对炎性痛症大鼠不同部位TRPV1的影响*
2020-08-01 07:45白艾灵赵征宇蔡定均
世界科学技术-中医药现代化 2020年5期
白艾灵,刘 祎,韦 婷,赵征宇,蔡定均
(成都中医药大学针灸推拿学院 成都 610075)
近年研究显示,TRPV1已然成为研究疼痛离子通道热点,而TRPV1(香草酸瞬时受体亚型1)能够在机体中感受热、酸以及内源性炎症因子等多种刺激,是炎性疼痛和机械性刺激的重要因子。众所周知,TRPV1是外周感觉神经元的伤害感受器,在急性和慢性炎症疼痛中皆具有重要地位,其可识别来自周围神经末梢的感觉输入,包括疼痛信号,随着TRPV1通道的激活从而诱导阳离子流入,并进一步激活信号传导途径[1-3]。有研究发现,背根神经节含有初级感觉神经元,负责接收对有害刺激反应的疼痛信号,并将信号传递给脊髓背角,在疼痛信号的整合、传导和外周敏化中发挥重要作用,如有研究表明,表达TRPV1的背根神经元,在通过疼痛、炎症等刺激而引发的信号转移中发挥关键作用[4-5]。
TRPV1 是感觉痛觉受体的重要信号整合因子,能够诱导敏化疼痛,若针刺干预则可通过一系列的应答反应来达到镇痛的目的[5-6]。大量研究证实,针灸具有显著的镇痛效果,且针刺镇痛呈现出明显的节律性[7-8]。背根神经节(代称DRG)和脊髓皆作为感觉传入神经元,其与针刺镇痛具有密切相关性[9]。针刺能缓解炎性疼痛,但由于针刺手法、穴位的选取和时间点不同,针刺对炎性动物的镇痛效果均不同。足三里穴在针刺镇痛研究中运用较为广泛,对炎症疼痛亦具有明显效应[10]。
本课题前期研究发现,以正常动物及弗氏炎症痛症等实验模型为载体,针灸的时辰差异可决定其抑制疼痛的疗效的不同,然而此间差异的机制如何,且该机制是否与TRPV1通道相关尚不清楚,故而该研究于前期实验的条件下,起于弗氏佐剂模型,并以TRPV1的表达作为效应指标,选取针刺镇痛效应的痛阈变化最优与最差两个点[11],研究选择不一样的时间点针刺足三里穴对弗氏炎症痛症大鼠中多个层次中TRPV1表达情况,去探索不同时间点针刺镇痛效应差异机制与TRPV1不同部位的蛋白表达的相关性。
1 材料
1.1 实验动物
雄性SD 大鼠(Wt:(160-230)g,四川省动物科研中心提供,SCXK(川)2013-15)。在成都中医药大学时间节律室内饲养,并使用CLOCKLAB 自动调控光照时间(早上七点开始光照,晚上七点关闭光照),通风1 h/1次,室温:(21±2)℃。
1.2 实验试剂
弗氏完全佐剂(以下简称CFA,F5881-10ML,101M8711,Sigma-Aldrich生物技术有限公司);生理盐水;TRPV1,兔多克隆抗体(ab63083,abcam 艾博抗贸易有限公司)。
2 方法
2.1 模型制备与实验分组
将所有实验大鼠于自动光暗周期驯养7 天后,采用辐射热甩尾法,先进行痛阈测定并筛选出实验对象,痛阈筛选标准为3-10 s 成功甩尾者即纳入为实验大鼠。然后再按照痛阈值随机分为待模型复制组(模型组)和生理盐水组(空白组)。模型组以1 ml 医用注射器在实验对象右足足底皮下注射0.1 ml CFA,空白组以同样手段在实验对象右足足垫部皮内注射等量生理盐水;24 小时后,观察大鼠右足情况以及其行为从而判断造模是否成果。将成功获得的模型实验鼠,依据其痛阈数值随机分为两个实验组,即模型组、模型针刺组(以下简称:模针组),每组按照不同时间点分别处理,又分为ZT8(15:00)、ZT16(23:00)两个时间组。
2.2 处理方式
模针组:于注射CFA 第二日分别在两个时间点,即ZT8 和ZT16,给予针刺,选穴足三里穴,共针刺25-35 min,每五分钟持续捻转手法行针1 分钟(频率:110-130 次/分钟),即每次针刺共行针5 次,而针刺处理时,需提前使用自制绑鼠工具捆绑老鼠,1日1次,连续治疗7日。
模型组、空白组的实验大鼠仅进行与模针组实验大鼠以相同自制固定器具并同样方式捆绑固定30 分钟处理。
2.3 检测指标
将从大鼠穴区皮肤、脊髓和DRG 中采集的组织样本运用免疫组化的方式测定目标物的表达情况,而免疫组织化学测定结果显示标志:阳性表达物、阴性表达和底色分别标记于不同的颜色即为浅黄色或棕黄色、蓝色、白色。使用Media Cybernetics 公司的Image-ProPlus 6.0图像分析检测采集的图像并确定其平均光密度。
2.5 统计方法
应用SPSS19.0 统计分析软件对所得实验数据以单因素方差统计方法分析,结果皆以平均数±标准差(xˉ±S)表示,以P<0.05认为差异具有统计学意义。
3 实验结果
3.1 ZT8时间点针刺调节大鼠不同层次中TRPV1的变化情况
ZT8时间点:①在穴区皮肤中,模型组大鼠皮肤样本中TRPV1的含量明显高于空白组大鼠中TRPV1的含量(P<0.05),而模针组可能由于针刺调节的影响其皮肤样本中TRPV1的表达明显低于模型组(P<0.05)。②大鼠脊髓中,模型组中脊髓节段样本中TRPV1的含量明显多于空白组(P<0.05);模针组脊髓样本组织中相较于模型组脊髓节段样本中TRPV1的表达明显下调了
表1 ZT8时针刺调节大鼠多个层次中TRPV1变化情况(± S;n=9)

表1 ZT8时针刺调节大鼠多个层次中TRPV1变化情况(± S;n=9)
注:与模型组相比,△P <0.05 △△P <0.01
背根神经节TRPV1 0.1523±0.0060△△0.1792±0.0093 0.1581±0.0060△△组别空白组模型组模针组皮肤中TRPV1 0.1729±0.0112△0.1877±0.0092 0.1729±0.0121△脊髓TRPV1 0.1401±0.0108△△0.1614±0.0106 0.1425±0.0111△△
表2 ZT16时针刺对大鼠各个层面组织中TRPV1调节情况(± S;n=9)
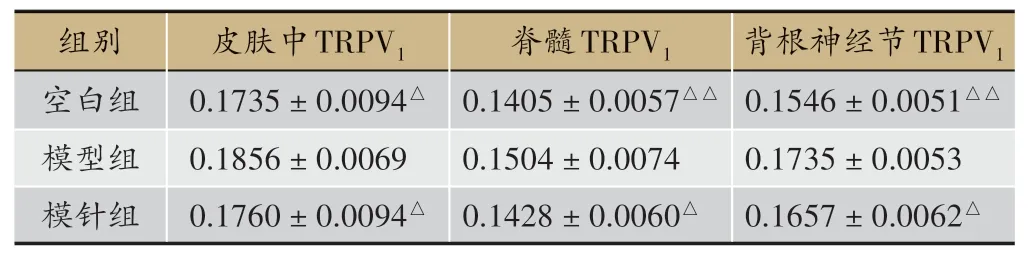
表2 ZT16时针刺对大鼠各个层面组织中TRPV1调节情况(± S;n=9)
注:与模型组相比,△P <0.05,△△P <0.01
背根神经节TRPV1 0.1546±0.0051△△0.1735±0.0053 0.1657±0.0062△组别空白组模型组模针组皮肤中TRPV1 0.1735±0.0094△0.1856±0.0069 0.1760±0.0094△脊髓TRPV1 0.1405±0.0057△△0.1504±0.0074 0.1428±0.0060△
TRPV1的表达量(P<0.05)。③在DRG 组织中,模型组DRG 中目标因子与空白组DRG 样本中TRPV1的增加量具有显著差异(P<0.01);模针组DRG 样本组织则比模型组DRG 中TRPV1表达量下调许多(P<0.05)。(见表1、图1、图2)
3.2 ZT16 时间点针刺调节大鼠各个部位中TPRV1表达的情况
ZT16 时间点:①穴区皮肤中,模型组与空白组比较,模型组大鼠皮肤样本组织中TRPV1的含量明显增加(P<0.05);模针组中大鼠皮肤TRPV1比模型组下调得更明显(P<0.05)②根据结果显示:在脊髓组织中,模型组大鼠脊髓节段样本中TRPV1的含量多于空白组(P<0.01);经过针刺处理后,即模针组大鼠脊髓节段中TRPV1的表达量明显减少(P<0.05)。③在DRG 组织TPRV1三个实验组中的变化情况与脊髓中TRPV1的表达情况类似,即模针组DRG 样本中目标物TRPV1的含量亦明显减少(P<0.05)。(见表2、图3、图4)
4 讨论
近年文献研究显示,针灸治疗痛症已经成为至关重要的手段之一。有研究发现,人类对于疼痛的感受亦具有一定的时间规律,但不同的病症表现出来的疼痛的阈值以及阈值最高值的时间亦具有差异性,如胆绞痛、偏头痛等疼痛阈值最高时大多发生于夜晚,然而某些关节炎性疼痛等疼痛阈值最高时间,大多亦可能发生于凌晨抑或是上午时段。有研究表明,于整个白昼与黑夜时段,晚上疼痛发作的次数较多,特别是上半夜甚至是整个昼夜的频发时段[12-13]。以上研究皆显示出痛症其本身即具有一定的节律,而且各种痛症所表现出来的节律还存在一定的差异性。

图1 ZT8时间点大鼠局部皮肤、脊髓及背根神经节TRPV1的表达(IHC 100×)

图2 ZT8时间点大鼠局部皮肤、脊髓及DRG中TRPV1的表达(IHC 200/400×)

图3 ZT16时间点大鼠局部皮肤、脊髓及DRG中TRPV1的表达(IHC 100×)

图4 ZT16时间点大鼠局部皮肤、脊髓及DRG中TRPV1的变化情况(IHC 200/400×)
相应地,在近年来的各种研究皆已经验证了时间子午流注法等中医传统的时间针灸疗法对于疼痛的治疗的明确效应,例如,有研究发现,采用时刻注穴法治疗疾病,结果显示其与对照组相比疗效具有统计学意义,且疼痛症状亦有明显改善[14]。现代基础研究亦提示了不同时间针刺在针刺镇痛中的重要地位,例如,在不同的间隔时间对佐剂性关节炎的大鼠进行针刺,针刺对于疼痛的效应具有明显的不同性,于间隔时间的不同进行针刺,结果显示在整个昼夜后针刺抑制炎症大鼠疼痛的疗效具有显著的提升,因此本研究提出针刺的时间跨距的差异所产生的抑制疼痛的效应的差异性,有可能与阿片肽基因密切相关[15]。有相关研究选用不同时间电针针刺,以此来观察外伤疼痛模型实验大鼠血浆褪黑素的浓度,在不同时间点针刺,结果显示正常大鼠血浆褪黑素浓度具有显著节律性,模针组治疗后血浆褪黑素浓度与模型组相比较,该浓度有一定的增加;而其节律性则和空白组类似,表明针刺可能会影响大鼠松果体分泌的褪黑素,且与外伤疼痛所引起的褪黑素节律紊乱的恢复具有密切关系[16]。因此针刺抑制疼痛的疗效确切,且具有一定的节律性。
疼痛具有保护作用,但其失调会对个人的生活产生负面影响,炎症性疼痛可由神经系统伤害感受器引起,且炎性化疼痛通常会引起神经纤维周围化学环境的高敏性。据研究显示,引发的炎性疼痛,通过动作电位、通道、受体和信号分子向上游传导至脊柱或皮质脑区,从而引起一系列的疼痛反应;炎性疼痛抑或由外周组织损伤或炎症引起,增加质子、组胺、三磷酸腺苷等的释放,这些物质的过度表达进一步导致外周和中枢致敏则引发疼痛[17-19]。疼痛的伤害感受器广泛分布于机体的各个组织,TRPV1作为对伤害性刺激有反应的重要通道,其在皮肤细胞中表达并充当疼痛或信号传感器;在炎症局部皮肤中释放的介质可能有助于外周致敏,特别是通过TRPV1致敏,从而促进疼痛的发生[20]。在脊髓及背根神经节的痛觉神经末梢和背角浅层均分布有TRPV1,其神经元感知外界有害刺激并传递信息到脊髓背角和三叉神经脊束核,从而在炎症痛症和神经源性痛症中起着至为关键作用[21]。总之,炎症疼痛通过各种信号通路来调节TRPV1的活性,以使得TRPV1的激活从而促进痛觉过敏的产生和维持。
有研究显示,针刺作为一种物理刺激,针刺刺激后可通过相应的神经传入中枢神经系统,引起不同的反应。脊髓为中枢神经系统的一部分,作为感觉传导的神经元,其在针刺镇痛的过程中起着不可或缺的作用[9,13]。背根神经节含有初级感觉神经元,负责接受对有害刺激反应的疼痛信号,从外周神经元传递的信号刺激脊髓伤害感受神经元,最后导致中枢敏化,其在这一过程中扮演着极为关键的角色[22]。研究者发现,在炎性疼痛过程中,TPRV1可以被P 物质等激活,再被PKC 磷酸化,然而电针针刺足三里穴可以通过影响某些物质的变化,从而达到抑制疼痛的目的23-24]。电针双侧足三里后,DRG 和脊髓中TRPV1水平皆显著减少,这表明针刺可以通过下调其TRPV1的过度表达或增加过多,来达到舒缓炎性痛症的目的。背根神经节中TRPV1水平亦与炎性疼痛的表达密切相关,以皮下注射完全弗氏佐剂来建立炎性疼痛模型,结果观察到在注射弗氏佐剂后第6天DRG中TPRV1的含量显著增加,然而电针后可以观察到其表达显著下调[25-26]。足三里穴区皮肤由脊髓神经节段所发出的神经支配,刺激足三里穴可以通过一定通路影响到脊髓背角的反应机制[27]。因此,针刺镇痛的机制不仅仅涉及单独的神经结构,而表明镇痛效应是由多层次、多水平的综合整合来反应的。
相应地,TRPV1是机械性刺激和热刺激的感受器,是研究疼痛机制中的特异性靶点,同时其与疼痛传导过程关系密切;有研究显示脊髓背角TRPV1表达的增加对初级中枢的功能具有致敏性[4,18,28]。有研究发现TRPV1和时钟基因Cry2 之间具有密切的关系,不仅如此,TRPV1的昼夜节律表达模式与昼夜节律系统之间具有紧密的相互作用[29-30]。该实验显示在ZT8 与ZT16时,炎性痛症大鼠中穴区皮肤、脊髓和DRG 中TRPV1的表达量皆明显增加或显著上调,表明炎性痛症可能诱发皮肤、脊髓和DRG 中TRPV1的上调;且其两个时间点大鼠穴区皮肤、脊髓和背根神经节中TRPV1的蛋白含量具有明显差异,而针刺足三里穴在ZT8、ZT16均显示此各个组织中目标因子TRPV1的表达明显下降,此亦进一步证明了针刺镇痛效应机制的复杂性、多样性,其不仅仅局限于某一区域,而是从外周到中枢的多层次、多水平的综合整合从而达到针刺镇痛的作用。
不仅如此,在ZT8、ZT16 针刺治疗炎性痛症大鼠时,大鼠痛阈均显示升高,但是其痛阈升高程度却具有明显差异性,而且其痛阈变化值在ZT8、ZT16 分别呈现出最优、最差两个方面,这也提示针刺时间不同,针刺镇痛效应具有一定的差异性亦可能与不同时间点各区域TRPV1 的蛋白含量差异相关。由上叙述可知,TRPV1与时间钟基因具有密切联系,其可能成为调控时钟基因的新参与者,因此,不同时间点出现的针刺镇痛效应的差异性可能与TRPV1本身昼夜节律的表达模式相关,但是其具体如何调控则并不清楚;而该实验从多个层面进行分析研究了不同时间点针刺镇痛效应差异与不同部位TRPV1表达的相关性,同时为临床针灸镇痛效应提供了非常有力的理论依据,因针对TRPV1可能的上下游信号分子机制亦未有清晰的研究分析,亦为后续研究或我团队亦正在针对该问题进行下一步的研究奠定了夯实的科学基础。
猜你喜欢
军事文摘(2022年8期)2022-11-03
中国现代医生(2022年21期)2022-08-22
中老年保健(2022年4期)2022-08-22
中国人兽共患病学报(2022年5期)2022-06-08
现代仪器与医疗(2022年1期)2022-04-19
散文诗世界(2022年2期)2022-03-10
昆明医科大学学报(2021年12期)2021-12-30
现代临床医学(2021年5期)2021-11-02
中国药学药品知识仓库(2021年18期)2021-02-28
电子制作(2019年7期)2019-04-25
