
基于零模型的宁夏荒漠草原优势种群点格局分析
2020-07-31 09:35许爱云许冬梅刘金龙黄葭悦米世明朱宁宁
生态学报 2020年12期
许爱云,许冬梅,2,*,刘金龙,于 双,黄葭悦,米世明,朱宁宁
1 宁夏大学农学院,银川 750021 2 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,银川 750021
植物种群空间格局分析是预测种群动态变化,揭示群落形成、维持与演替内在机制及生态过程的重要手段[1-2],植物看似无序的分布格局,其潜在的生态过程和形成机制实质是环境资源分布形式与组合方式的反馈[3]。种群空间分布格局是多种生态过程综合作用的结果[2],只是在不同时空尺度与群落类型中,各自所起的作用强度有所不同[4- 5]。明确植物优势种群个体的空间分布格局可以反映种群的生态适应对策,揭示优势种群空间格局的形成机制有助于解释群落物种共存与多样性维持的机制[1,6]。
植物种群间相互作用和生态过程所发生的空间区域是生态动力学的一个重要决定因素,利用空间数据基于生态过程的理论假设来模拟验证生态理论是生态学研究的内容之一[7-9]。由于点格局分析法以植物个体在二维空间上的坐标为基础,极大限度的利用了植物种群空间坐标图的信息[10],能够量化描述不同尺度范围内种群的分布特征,多尺度全面反映植物种内种间空间格局信息,检验能力较强[6,9],因而被广泛应用于植物种群分布格局的研究中。
基于生态位理论的生境过滤与基于中性理论的扩散限制能较好地解释种群的环境适应性和资源利用能力[4]。关于生境过滤与扩散限制机制分别在物种多样性[11]、土壤微生物群落构建[12]与物种空间分布格局[2,13-14]等研究中得到广泛的应用和较好的验证,但这些研究多是基于局地尺度对森林生态系统的研究,较少涉及草地生态系统,尤其是荒漠草原生态系统。宁夏荒漠草原地处宁夏中北部,风力作用强盛,风沙地貌发育,是生态环境脆弱敏感的区域[15]。近年来随着全球气候的变化,加之人类活动的过度干扰,导致荒漠草原区植被退化,生产力降低,植物群落呈现出异质性较高的空间分布格局[15-17]。为进一步了解宁夏荒漠草原优势种群的空间分布格局及主要驱动机制。本文基于不同零模型的点格局分析方法,以宁夏荒漠草原优势种群蒙古冰草(AgropyronmongolicumKeng.)、短花针茅(StipabrevifloraGriseb.)、牛枝子(LespedezapotaniniiVass.)和牛心朴子(CynanchumkomaroviiAL.lljinski.)为研究对象,在分析其种群空间分布格局特征的基础上,讨论生境异质性、扩散限制等驱动因子在其空间分布格局形成过程中的作用,旨在揭示严酷的荒漠草原生境中优势种群空间格局的分布规律及其对环境的适应特征,以期为退化荒漠草原的恢复与重建提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于宁夏东部盐池县花马池镇四墩子村(107°15′40″E,37°20′12″N),地处毛乌素沙地南缘,黄土高原向鄂尔多斯台地的过渡地带(图1)。自然条件较为恶劣,干旱少雨,风沙大,属典型的中温带大陆性季风气候[15],年平均温7.7 ℃,1月和7月平均气温分别为-8.9 ℃和22.5 ℃,年温差较大,≥0 ℃积温为3430.3 ℃;年平均降水量289.4 mm,主要集中在6—8月份,年蒸发量为2135 mm,年无霜期165 d。土壤类型主要为灰钙土、风沙土,土壤质地为沙壤和粉沙壤,土壤结构松散,肥力低下[18]。植被类型属荒漠草原,主要植物种有:蒙古冰草(Agropyronmongolicum)、牛枝子(Lespedezapotaninii)、短花针茅(Stipabreviflora)、牛心朴子(Cynanchumkomarovii)、黑沙蒿(ArtemisiaordosicaKrasch.)、赖草(LeymussecalinusTzvel.)、甘草(GlycyrrhizauralensisFisch.)、猪毛蒿(ArtemisiascopariaWaldst. et Kit.)等。
图1 研究区地理位置Fig.1 Location of the research area
1.2 样地设置与取样
于2018年7月在宁夏大学四墩子教学科研基地所处的荒漠草原区,选取、设置地表平坦,分别以蒙古冰草(Plot 1)、短花针茅(Plot 2)、牛枝子(Plot 3)和牛心朴子(Plot 4)为优势种群的10 m×10 m的典型样地共4个。在每个样地,采用邻接格子法,用1 m×1 m的样方框按从东到西,从南到北的顺序依次放置100次进行取样,以样方框一角顶点为基点(0,0),确定、记录各优势种群个体在样地内的二维坐标。最后,在室内对取样记录进行处理,按照取样时样方框放置的顺序将100个1 m×1 m的样方图进行拼接,合并成10 m×10 m的样地图,用于点格局分析[19]。各样地概况见表1,各种群个体分布位点见图2。
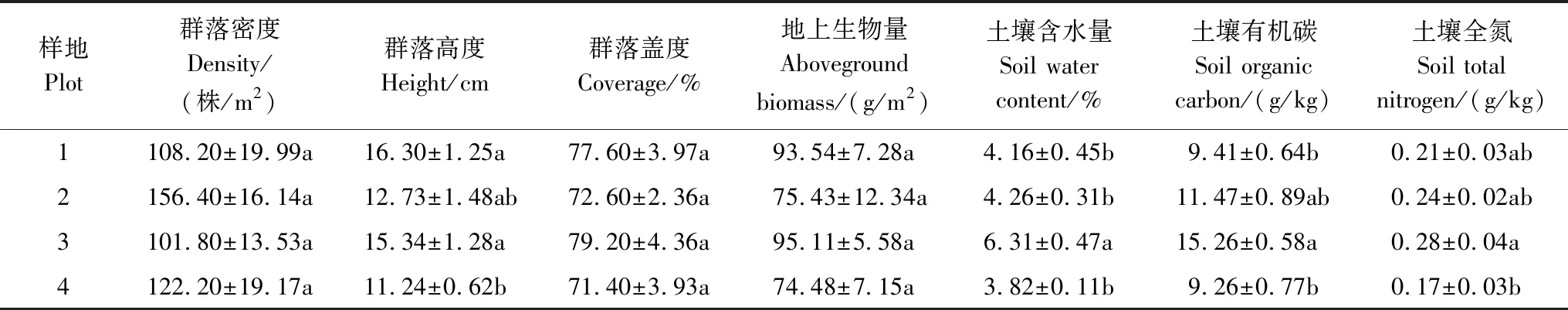
表1 不同研究样地概况

图2 不同种群个体分布点图Fig.2 Point diagram of individual distribution for different populations
1.3 分析方法
1.3.1 点格局分析
点格局分析由Ripley[20]提出,分析连续尺度上种群格局的特征,是目前植物生态学领域研究种群空间格局广泛应用的方法之一[7-8]。其中,O-ring函数是基于Ripley′s K修正的函数,克服了K(r)函数随尺度累计效应的不足,使得种群格局的分析结果更准确[6- 7],具体见宁夏荒漠草原蒙古冰种群小尺度空间分布及其关联性研究[21]。
在实践中,通过选定零模型(null model)与O(r)函数相结合,利用Monte-Carlo随机模拟,并利用模拟的最大值和最小值生成上下两条包迹线,即置信区间来分析种群空间格局[19]。本项研究中,选择O(r)函数,并根据样方大小设圆环宽度为1 m,检验尺度为0—5 m,利用 Monte-Carlo随机模拟99次,拟合检验上、下包迹线,即99%的置信区间[2,21]。
1.3.2 零模型选择
空间格局复杂多样,选择合适的零模型对空间点格局分析尤为重要,不同的问题假设和研究目的需要选择不同的零模型,为了解宁夏荒漠草原优势种群的空间分布格局,检验生境异质性与扩散限制对其空间格局形成的作用,本研究选取完全空间随机(Complete Spatial Randomness, CSR)、异质泊松(Heterogeneous Poisson, HP)和泊松聚块(Homogeneous Thomas, HT) 3种零假设模型[13,19,22]。完全空间随机模型实质是同质泊松过程,该模型假设中物种个体的空间分布不受任何生物或非生物过程影响,是相互独立的,即在研究区域内任何位置上出现的概率相同,是判断种群空间分布格局的主要零模型[19,23]。而异质泊松模型与泊松聚块模型分别用于检验生境异质性与扩散限制是否影响种群空间分布格局,它们是在CSR模型应用基础上,通过排除生境异质性或扩散限制因子进一步模拟验证[2,13,24]。该模拟过程中,如果O(r)的实测值落在置信区间,没有发生偏离,物种呈随机分布,则该模型假设的生态过程对该物种空间分布有显著影响[13];如果O(r)的实测值高于或低于置信区间,发生偏离,则说明该模型假设的生态过程不能完全或有效地解释所研究物种空间分布格局形成的过程[23]。
1.4 数据处理
采用Programita 2014软件进行圆环函数O(r)不同零模型的点格局计算,Origin 8.0进行绘图。其他数据处理采用Excel 2013软件完成。
2 结果与分析
2.1 基于完全空间随机模型的点格局分析
完全空间随机模型的点格局分析表明(图3),蒙古冰草种群和牛心朴子种群O(r)实测值分别在0—3.3 m和0—0.5 m尺度范围位于置信区间之上,表现为聚集分布;随着尺度的增大,O(r)实测值位于或低于置信区间,种群逐渐过渡到随机分布和均匀分布。短花针茅种群和牛枝子种群分别在0—3.6 m和0—3.3 m尺度范围内位于置信区间之上,表现为聚集分布,而在其余尺度均服从随机分布。可以看出,荒漠草原区优势植物种群的空间分布格局在小尺度(<4 m)上主要表现为聚集分布,随着尺度的增大,聚集程度减弱,逐渐过渡到随机分布和均匀分布。
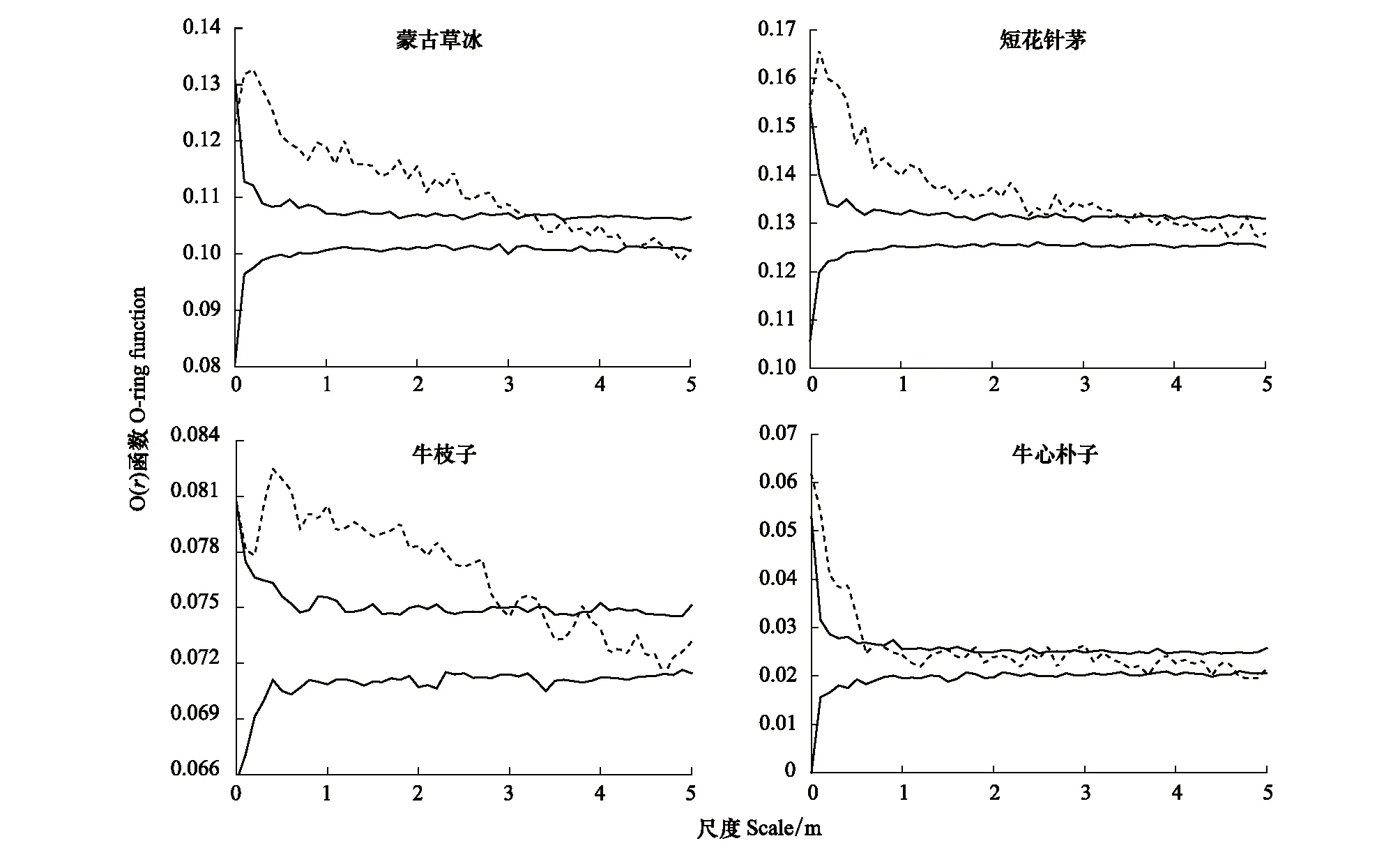
图3 基于完全空间随机模型的优势种群点格局分析Fig.3 Point pattern analysis of dominant populations based on complete spatial randomness model
2.2 基于异质泊松模型的点格局分析
基于排除生境异质性影响的异质泊松模型分析显示(图4),蒙古冰草种群在整个研究尺度内符合异质泊松模型,表现为随机分布;短花针茅种群与牛心朴子种群仅分别在0.1—0.4 m、0—0.2 m小尺度范围偏离异质泊松模型,表现为聚集分布,在其他尺度范围均表现为随机分布;牛枝子种群除了在0—0.2 m小尺度范围偏离异质泊松模型,表现为均匀分布,其他尺度范围均表现为随机分布。
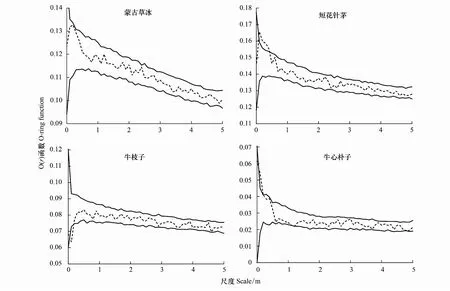
图4 基于异质泊松模型的优势种群点格局分析Fig.4 Point pattern analysis of dominant populations based on heterogeneous poisson process model
2.3 基于泊松聚块模型的点格局分析
基于泊松聚块模型剔除扩散限制影响后,发现蒙古冰草、短花针茅、牛枝子和牛心朴子4个种群在所研究尺度范围内均没有发生偏离,符合泊松聚块零假设模型,表现为随机分布(图5)。与异质泊松零模型的检验结果相比,该模型假设的扩散限制机制较好的解释了荒漠草原优势种群空间格局形成的过程。

图5 基于泊松聚块模型的优势种群点格局分析Fig.5 Point pattern analysis of dominant populations based on poisson cluster process model
3 讨论与结论
种群空间分布的聚集程度和尺度转化可体现物种对生境的适应,资源的利用以及物种的繁殖特性[23]。在完全空间随机模型下,宁夏荒漠草原4个优势种群在不超过4 m的尺度范围内主要表现为聚集分布,其中蒙古冰草、短花针茅种群的偏离程度较高,且聚集尺度也较大,其次是牛枝子种群,偏离程度最低、聚集尺度最小的是牛心朴子种群。该结果与蒙荣[25]和王鑫厅[26]等人的研究结果相似,符合自然群落种群聚集分布的一般规律[27]。种群在一定尺度内表现出聚集分布特征,有利于其发挥群体效应,增强抵御外来危害的能力,维持种群正常发展[27-28]。本研究区地处毛乌素沙地南缘,风蚀作用强烈,种群个体抵御风沙危害的能力有限,为了适应较为严酷的自然生境,提高存活机会,种群内个体间彼此相互合作,相互庇护,进而表现出聚集分布的特征。除了受生物学特性影响,这种聚集分布与偏离程度的差异可能与生境异质性或扩散限制有关。
在干旱、半干旱草原生态系统中,地境因子较高的空间变异是一个普遍的特征[29],异质生境为物种共存提供了环境资源,对群落结构、物种多样性与种群的动态变化具有重要的影响[4,11,30]。Getzin等[31]通过研究同质与异质生境中铁杉种群的空间格局,证实生境异质性在植物种群结构与空间格局形成中具有重要的作用。本研究在完全空间随机模型下,各优势种群在小于4 m尺度范围均呈明显的聚集分布,但采用异质泊松零模型,消除生境异质性后,蒙古冰草种群呈随机分布;短花针茅、牛枝子和牛心朴子种群仅在0—0.4 m尺度范围发生偏离,其他尺度均呈随机分布,说明荒漠草原优势种群空间分布格局形成中生境异质性起到重要作用。由于小尺度内空间连续性较好,环境变量相对均质,导致生境异质性的作用降低[4],使短花针茅、牛枝子和牛心朴子种群在0—0.4 m尺度范围发生偏离。其他研究也显示在小尺度范围内生境异质性仅对部分种群的分布有显著影响[2,22,30],而在大尺度上(>10 m),环境异质性被认为是影响种群空间分布的主要因素[22,31]。另外,该结果也可以间接反映各优势种群对生境的偏好程度,蒙古冰草种群在排除生境异质性后,完全符合异质泊松模型,呈随机分布,可见它具有独特的生境依赖性或生物学特性[32]。
种子扩散是种群动态中极为重要的生态过程,也是解释种群空间格局变化的重要机制[33-34]。种群的空间聚集强度、尺度与种子的扩散方式有关[35]。种子扩散不仅受风力、水力、动物及地形等环境因素的影响,而且与植物的繁殖特性和生物学特性密切相关[36-37]。本研究在基于完全随机零模型假设基础上,通过泊松聚块零模型排除扩散限制后,所研究种群均没有发生偏离,完全符合泊松聚块零模型,呈随机分布。说明在宁夏荒漠草原,种子扩散限制也是导致优势种群分布格局形成的重要机制。这可能与种群的生殖策略有关,研究表明,短花针茅在环境恶劣的条件下,有性生殖产生的个体竞争能力减弱,而靠基部分蘖产生个体的营养繁殖成为其有限范围内扩展空间的主要方式[38]。牛枝子匍匐状的生长特征,使其种子散落后主要集中在地表的局部范围,导致其种群在小尺度范围内常呈聚集分布[39]。随着尺度的增大,种内密度制约效应加剧,种子扩散和繁殖能力减弱,种群逐渐趋于随机分布或均匀分布。此外,在干旱草地生态系统中,植被也是影响种子扩散的因素之一[40- 42]。本研究中,4个群落样地植被盖度均较高,一定程度上限制了种子的扩散。
综上,宁夏荒漠草原优势种群的空间分布格局在小于4 m尺度范围呈聚集分布,生境异质性和扩散限制均是驱动其空间分布格局形成的重要机制。相对而言,异质泊松零模型下,各优势种群符合模型的程度不及泊松聚块模型,即小尺度范围内(<0.4 m),生境异质性不能成为解释优势种群空间格局形成的理想机制。此外,研究尺度、种群竞争、密度制约、邻体效应、群落结构及其交互作用均会影响种群的空间分布[2,13,23]。因此,本研究结果还不能完全揭示各种群潜在信息,故结合种群组成与发育特征选择更多的零模型或不同因子交互作用的模型模拟验证来揭示种空间群格局的形成机制仍需进一步研究。
猜你喜欢
数学物理学报(2022年5期)2022-10-09
数学物理学报(2021年6期)2021-12-21
北方论丛(2021年2期)2021-05-22
落叶果树(2021年6期)2021-02-12
数学物理学报(2019年4期)2019-10-10
现代园艺(2018年1期)2018-03-15
体育科技(2016年2期)2016-02-28
云南师范大学学报(自然科学版)(2015年5期)2015-12-26
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
