
短柄枹栎林优势种群空间结构分析
2020-07-27 06:07:26彭舜磊张志录李彦娇
森林与环境学报 2020年4期
彭舜磊,齐 光,张志录,李彦娇,陈 栋
(1.平顶山学院化学与环境工程学院,河南平顶山467000;2.信阳师范学院数学与统计学院,河南信阳464000)
种群空间结构分析对了解种群的形成机制和种间关系具有重要意义[1]。很多学者对森林优势种群空间分布格局进行了分析,普遍发现优势种群和特有种群在一定尺度内多呈现聚集分布的格局[2-5],非优势种和特有种的空间分布格局多为随机分布或均匀分布,这种空间格局的形成有利于森林群落各类种群的自我发展及与其它种群的共存[6-8]。种群空间分布格局随尺度变化而变化,不同生境和林型的优势树种的空间分布格局和种群结构差异很大[9-14];此外,森林内不同径级优势树种空间分布格局存在不同[1,11,14],优势种群径级结构不但反映竞争关系,而且反映种群大小结构[15-18]。生境的异质性和种群自身的生物学特性以及竞争关系影响种群的空间分布格局和径级结构[15-17]。
在宝天曼自然保护区,短柄枹栎[Quercus serrata var.brevipetiolata(A.DC.)Nakai.,Qs]林分布在海拔1 300 m左右的中山地带,是该保护区重要林型之一。藏亚静等[18]发现该林区1 hm2样地内,短柄枹栎和四照花(Dendrobenthamia japonica var.chinensis Osborn.,Dj)等7个生活型物种的空间分布均为聚集分布;沙迎迎等[19]研究表明,短柄枹栎和四照花等为聚集分布,化香树(Platycarya strobilacea Siebold et Zucc.,Ps)为随机分布。但是由于短柄枹栎林在宝天曼自然保护区内呈片段化分布,不同样地短柄枹栎林内优势树种的空间结构是否存在异质性,尚待深入研究。通过研究短柄枹栎林典型样地内优势树种的空间分布格局和径级结构,揭示其优势树种的空间结构特征,为宝天曼自然保护区短柄枹栎林生态恢复与可持续经营提供理论依据和科技支撑。
1 材料与方法
研究区位于河南省南阳市内乡县宝天曼自然保护区(111°53′~112°04′E, 33°25′~33°33′N)。 该区属暖温带向北亚热带过渡区,大陆性季风气候,年平均气温15.1℃,年平均降雨量885.6 mm,年平均蒸发量991.6 mm[18-19]。地貌以中山为主,最高海拔1 830 m;植被类型以暖温带落叶阔叶林为主,兼有南北过渡植物区系,短柄枹栎林主要分布在海拔1 100~1 300 m[18-19]。
1.1 样地设置
2014 年 8 月, 在宝天曼自然保护区海拔 1 327 m 处(111°55′41.1″E, 33°30′19.4″N), 选择短柄枹栎林分布的典型地段,设置1块20 m×50 m样地。该样地林龄为80 a,坡度38°,坡向南偏西15°,郁闭度0.85。将样地分为10个10 m×10 m的小样方,以样地的西南角为坐标原点,东西方向为x轴,南北方向为y轴,建立直角坐标系,记录每个树种的物种名和坐标,测定每个树种的胸径和树高。
1.2 数据分析
1.2.1 重要值 通过相对多度、相对频度和相对优势度计算重要值[19],利用重要值判定短柄枹栎林的优势树种,计算公式如下:

1.2.2 点格局分析方法 该方法以样地内树木的坐标点形成的空间分布点图为基础,用Ripley′s K函数分析树木的空间分布格局[20]。Ripley′s K函数是研究样地内以某点为圆心,以一定长度r为半径的圆内点数(植物个体数目)的函数,计算公式如下:

式中:K(r)为从某一随意点起距离r以内点数的期望;A为样地面积(m2);n为样地内树木个体总数(即总点数); Uij为点 i和点 j之间的距离(m), 当 Uij≤r时, Ir(Uij)= 1, 当 Uij>r时, Ir(Uij)= 0; Wij为权重值[20-21]。
采用完全随机模型,用蒙特卡洛方法模拟99次,得到上下包迹线围成的99%的置信区间,K(r)值在置信区间之上,种群为聚集分布;K(r)在置信区间之下,种群呈均匀分布;K(r)在置信区间之内,种群为随机分布[2,11,20-21]。用R 3.5.1软件中spatstat程序包的envelope函数做点格局分析,用ggplot 2程序包绘制样地树种空间位置图,用Excel 2016软件绘制径级结构图。
1.2.3 径级结构分析 将短柄枹栎林内乔木物种的胸径划分为0~5 cm、5~10 cm、10~15 cm、15~20 cm、20~25 cm、25~30 cm、30~35 cm和>35 cm八个径级,统计所有树种以及优势种每个径级对应的个体数,画出径级结构分布图,分析种群的径级结构。
2 结果与分析
2.1 群落物种组成
短柄枹栎林样地中乔木层内共有11种乔木物种,分别为短柄枹栎、华榛(Corylus chinensis Franch.,Cch)、化香树、四照花、千金榆(CarpinuscordataBl.,Cc)、三桠乌药(Lindera obtusiloba Bl.,Lo)、棠梨(PyrusbetulifoliaBunge, Pb)、 玉玲花(StyraxobassiaSieb.Zucc, So)、 栓皮栎(QuercusvariabilisBl.,Qv)、鹅耳枥(CarpinusturczaninowiiHance,Ct)、青皮槭(AcercappadocicumFranch.,Ac)。由表1可知,短柄枹栎、华榛、化香树、四照花重要值分别为29.46、22.65、15.41和11.98;华榛相对优势度最大(64.49%),相对多度最低(0.97%),研究该优势树种空间格局没有意义。因此,仅把短柄枹栎、化香树和四照花作为优势树种,研究其空间结构特征。
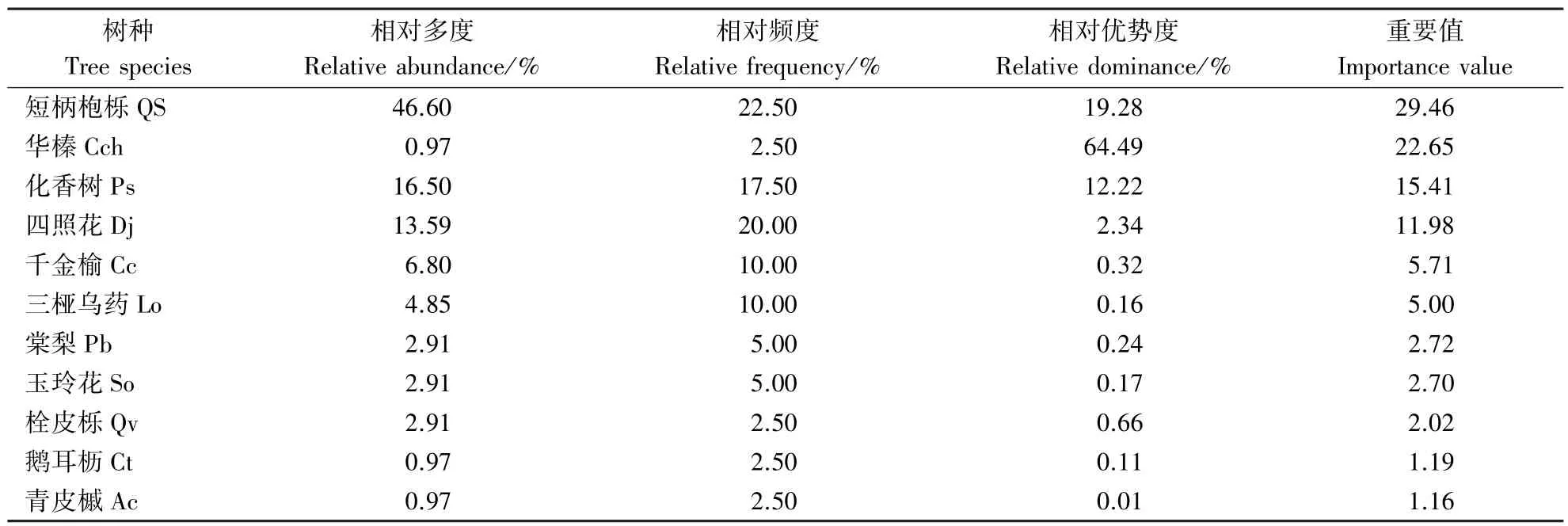
表1 短柄枹栎林乔木层树种的重要性分析Table 1 Analysis of the importance values of the tree species in Qs forest
2.2 优势种群的空间结构
图1表明,短柄枹栎林乔木层树种的个体分布很不均匀,成簇、成块的密集分布。短柄枹栎、四照花、化香树等优势种群的大小、群间的距离、群内个体的密度等都不相等。短柄枹栎种群分布范围较广,每棵大树周围都有小树分布,呈现集群分布。同种群的大树基本上都分布在一起,呈集群状态,而小树则是伴随在大树周围,分布很不均匀。
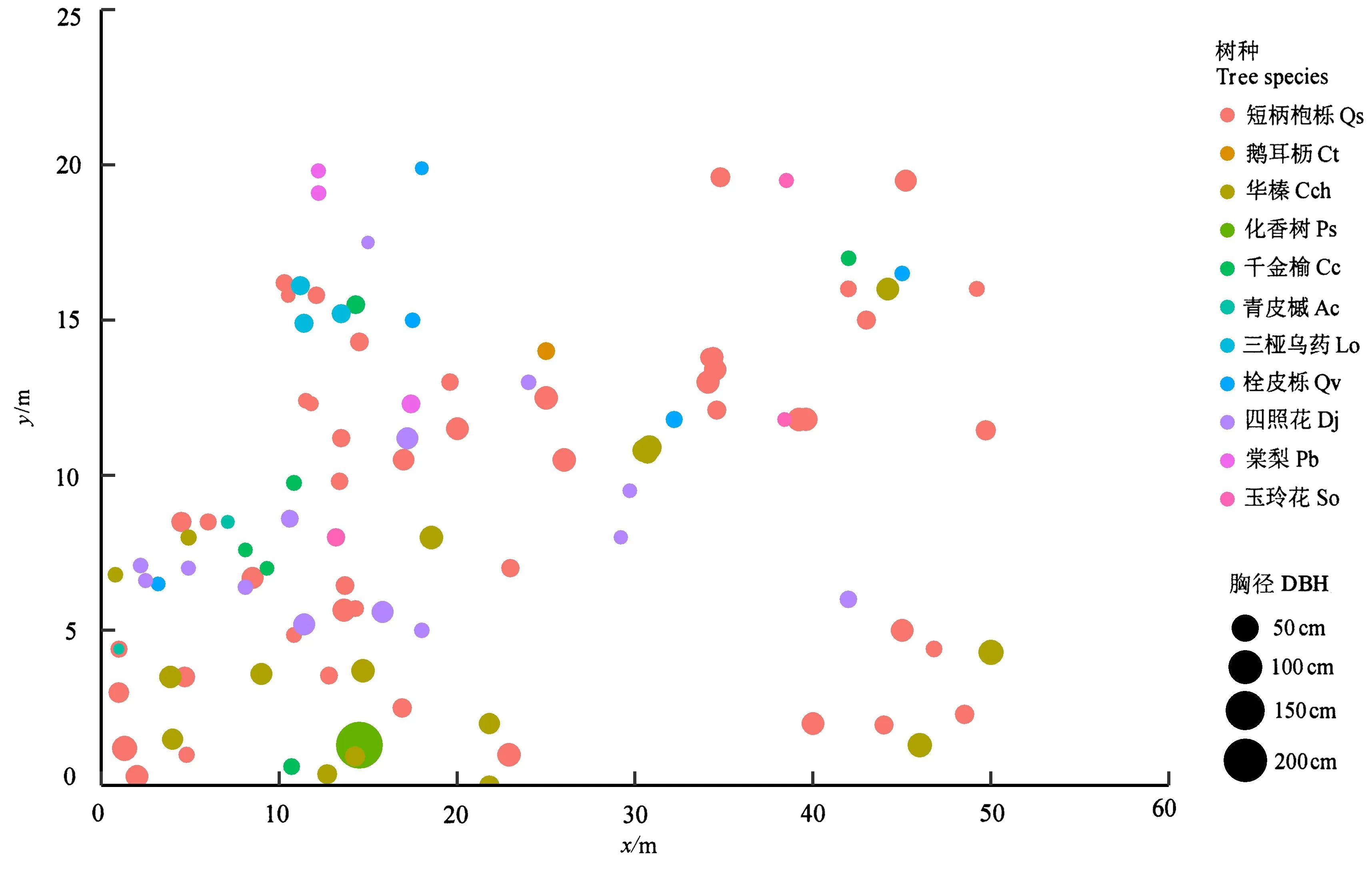
图1 短柄枹栎林样地内所有树种空间位置分布Figure 1 Spatial location of all tree species in the sample plot of Qs forest
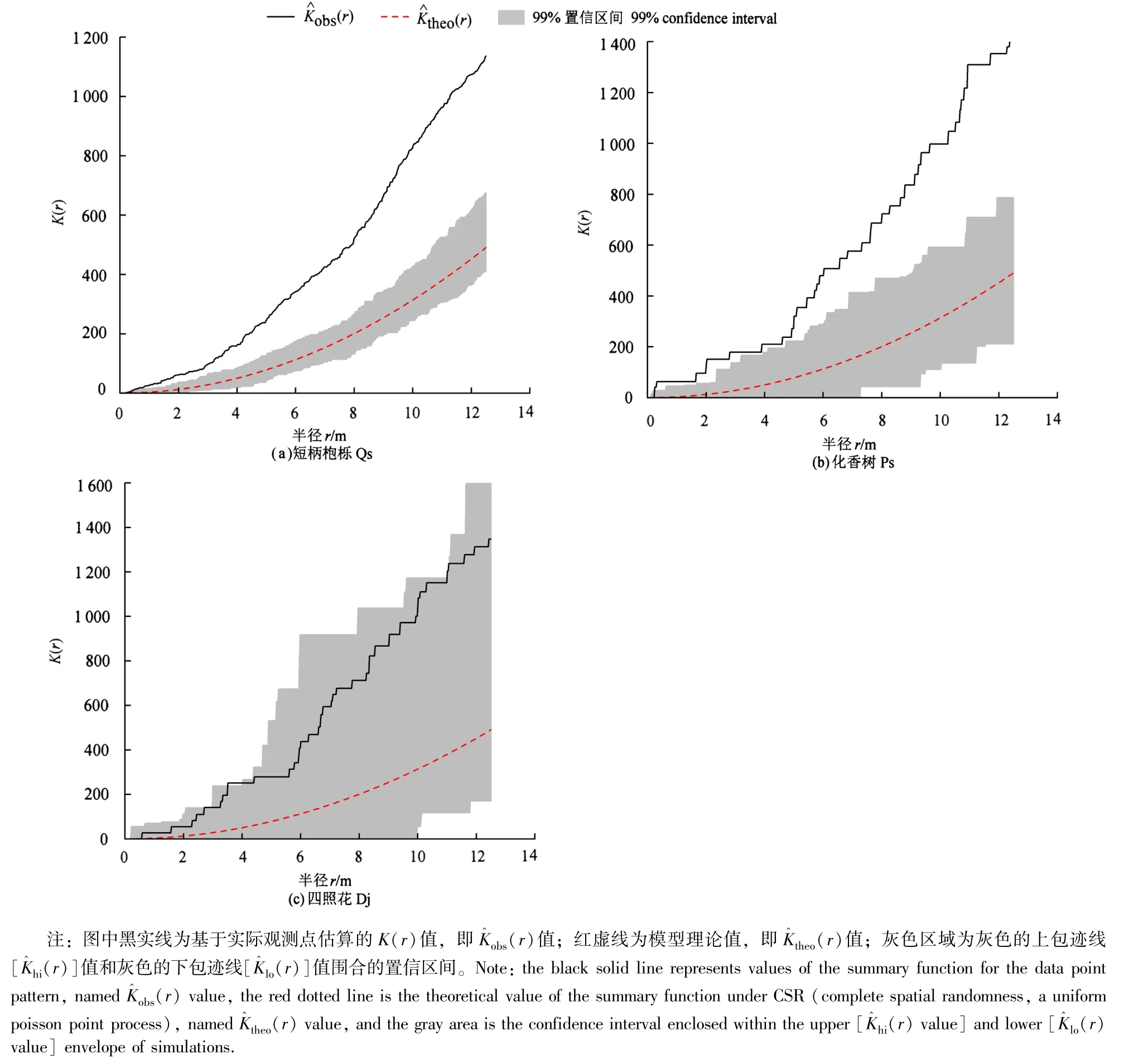
图2 短柄枹栎林优势种群空间分布格局Figure 2 Spatial distribution pattern of the dominant tree species in Qs forest
2.3 优势种群的径级结构
由图3(a)可看出,乔木层所有树种的径级结构呈近倒 “J”字型,年幼个体最多,占总个体数量的42.7%,而大树占7.8%,说明该林分整体为增长型。图3(b)表明,短柄枹栎的径级结构呈近正态分布,0~10 cm径级,个体数量随着径级的增加而增加,>10 cm径级,除25~30 cm径级个体数量较高外,其余径级的个体数量随着径级增加而逐渐减小,整体来看,该优势种为稳定型;小径级个体数量占该物种总个体数量的31.3%,中径级占64.6%,大径级占4.2%。由图3(c)可知,化香树的径级结构呈近 “J”型趋势,缺少0~5 cm和10~15 cm径级,该优势种小径级个体数量占总个体数量的11.8%,而中、大径级占88.2%,说明该优势种可更新的幼苗较少,种群呈现出衰退的趋势。由图3(d)可知,四照花的径级结构整体上呈近倒 “J”型趋势,在整个径级范围内,种群个体数量随着径级的增加而减少;径级为0~10 cm的树木株数最多,占总数的71.4%,说明该优势种正处于生长发育阶段,属于增长型,但所缺径级较多,不稳定。

图3 短柄枹栎林所有树种和优势树种径级结构分布Figure 3 Diameter class structure distributions of all tree species and the dominant tree species in Qs forest
4 讨论与结论
依据重要值,短柄枹栎、华榛、化香树和四照花4个树种的重要值较大,分别为29.46、22.65、15.41和11.98。剔除出现频度较小的华榛,短柄枹栎、化香树和四照花为该样地的优势物种。短柄枹栎林内短柄枹栎和化香树种群的空间分布格局为集群分布,而四照花种群的为明显的随机分布,与臧亚静等[18]和沙迎迎等[19]的研究结果并完全不一致,是样地所处的地段和生境不同导致的差别。植物种群空间分布格局受植物生物学特性(种子扩散和繁殖、密度制约、种内和种间竞争等)与生境因子(地形、土壤、雨量、光照等)共同影响[8,22-26]。由于短柄枹栎和化香树的种子大,主要散落在母树的周围,传播受限,易形成聚集分布[6-10]。种群呈聚集分布是一种好的趋势,在群落演替中后期,多个种群呈聚集分布,是竞争后的结果,优势树种短柄枹栎和化香树呈聚集分布,各自都有各自的生存空间,把其余的空间留给其它种群,有利于群落物种多样性维持[1-2,13]。四照花的生活型为小乔木,在林内竞争力弱,因此,四照花种群容易产生随机分布的空间格局。
短柄枹栎林样地内所有树种的径级结构呈近倒 “J”字型,说明乔木层树种的整体发展动态为增长型。短柄枹栎种群的径级结构呈近似正态分布,主要以中径木分布为主,幼树个体数相对较少,与沙迎迎等[19]的研究结果一致。化香树种群的径级结构呈近 “J”型,主要以大径级树木分布为主,幼树数量少,种群呈现出衰退的趋势。四照花种群的径级结构为近倒 “J”型,种群呈现出增长趋势。短柄枹栎林内化香树种群缺乏可更新的幼苗,承受着较大的竞争压力[11-14],原因是短柄枹栎林郁闭度大,光照不充足,不利于林下种子萌发和幼苗更新[18-19],而四照花幼苗充足,表明该物种生态位窄,幼苗适应林下环境能力强,表现出增长的趋势。短柄枹栎、化香树都是喜光物种,林下更新困难,建议采取适当的择伐等营林措施,促进林下更新。
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
核农学报(2020年6期)2020-06-30 06:15:38
植物研究(2018年4期)2018-07-24 01:06:24
西北植物学报(2015年9期)2015-07-05 07:03:42
河池学院学报(2014年5期)2014-02-27 13:37:18
