
舟山沿岸渔场春秋季鱼类群落结构特征分析*
2020-07-26 09:45郭朋军江新琴俞存根邓小艳许永久
渔业科学进展 2020年4期
郭朋军 江新琴 俞存根 邓小艳 张 平 许永久
舟山沿岸渔场春秋季鱼类群落结构特征分析*
郭朋军1,2江新琴2俞存根2①邓小艳2张 平2许永久2
(1. 舟山市海洋与渔业局 舟山 316000;2. 浙江海洋大学 舟山 316022)
根据2015年11月(秋季)、2016年5月(春季)在舟山沿岸渔场开展的渔业资源调查中所获得的鱼类资料,采用群聚的物种相似性、多样性指数、RDA分析等方法,对该海域鱼类种类组成、生态类群、多样性和群聚特征进行了分析。结果显示,本调查共鉴定出鱼类81种,隶属于12目39科63属,以近海暖水种为主;Jaccard相似性指数为37.04%,处于中等不相似水平;优势种计算结果显示,春季优势种为六丝钝尾虾虎鱼(Amblychaeturichthys hexanema),渔获量占总渔获量的42.16%;秋季优势种为龙头鱼(Harpodon nehereus),占总渔获量的43.56%;春季多样性高于秋季,但2个季节的多样性指数平面分布相似,均为调查海域南部最低,越往北多样性指数越高,且东部海域高于西部海域;ANOSIM检验显示,各群落间差异极显著;相似性百分比(SIMPER)显示,蓝圆鲹(Decapterus maruadsi)、棘头梅童鱼(Collichthys lucidus)、龙头鱼等7种,中华小沙丁鱼(Sardinella nymphaea)、鳀(Engraulis japonicus)、七星底灯鱼(Benthosema pterotum)等9种,分别是春、秋季各组群的典型种及造成鱼类群落结构差异的主要分歧种。RDA研究结果显示,盐度、温度是舟山沿岸渔场鱼类群落结构主要的环境影响因子。
鱼类;种类组成;多样性;群落结构;舟山沿岸渔场
鱼类是海洋生态系统的重要组成部分,其群落结构变化在一定程度上能够反映出生态系统的物质转换和能量流动(单秀娟等, 2014)。目前,对中国海域鱼类群落结构的研究已有很多(林楠等, 2009; 李圣法等, 2005、2007; 沈金鏊等, 1987)。舟山渔场作为中国海域的重要组成部分,对舟山渔场鱼类群落结构研究主要集中于对某一海域或邻近海域的研究(陈佳杰, 2012; 卢占晖等, 2009; 张洪亮等, 2012; 梁君等, 2014; 钟毅飞等, 2016),对整个舟山沿岸渔场鱼类群落结构详细调查的研究较少。受长江、钱塘江等大型淡水河流的径流作用,以及沿岸流、台湾暖流和黄海冷水团交汇作用,舟山沿岸渔场是许多鱼类重要的产卵场和索饵场(汪振华等, 2011; 程郡等, 2014)。因此,在舟山沿岸渔场开展资源调查、摸清该海域资源现状,对该海域渔业资源保护以及生态文明建设具有重要意义。本研究根据2015年11月和2016年5月在舟山沿岸渔场调查的鱼类资料,研究该海域鱼类种类组成、生态类群、优势种及群落格局,以期为舟山沿岸渔场鱼类资源的保护和合理开发利用提供科学依据。
1 材料与方法
1.1 数据来源
本研究所用数据取自2015年11月(秋季)、2016年5月(春季)“浙普渔32207号”单拖网船在舟山沿岸渔场29°20′~31°00′N、121°40′~123°00′E范围内实施的渔业资源调查时所获得的鱼类资源资料,在该范围内共设80个调查站位(图1)。采样网具为底拖网,网具规格为800目×80 mm,每个站位拖曳时间约为1 h,平均拖速约为3.5 kn。放网后,使用CTD仪对调查站位的温度、盐度等环境因子进行测定。记录渔获物中鱼类的网产量,统计每个物种的重量和尾数,并对每个物种进行生物学测定。

图1 渔业资源调查站位
1.2 数据分析
1.2.1 优势种 鱼类优势种的计算采用相对重要性指数,计算公式如下:
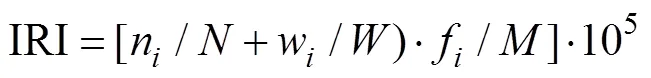
式中,n、w分别为第种生物的个体数和生物量;分别为调查所获得总个体数和总生物量;f为第种生物在次取样中出现的频率;为取样次数。将相对重要性指数(IRI)值大于1000者定为优势种,数值在100~1000之间者定为常见种(曹东阳等, 2015)。
1.2.2 生态类群划分 以《中国海洋鱼类》(陈大刚等, 2015)、徐兆礼(2008、2011)等为参考,按鱼类适温性及适盐性对舟山沿岸渔场鱼类划分生态类群。
1.2.3 群聚的物种相似性 采用Jaccard相似性系数(孙莎莎等, 2013)计算春秋季鱼类群聚物种组成的相似性。计算公式如下:

式中,为春季的种类数,为秋季的种类数,AB为春季和秋季共有种类数。当C为0~25%时,为极不相似;25%~50%为中等不相似;50%~75%为中等相似;75%~100%为极相似(李捷等, 2010)。
1.2.4 多样性分析 多样性指数采用Shannon- Wiener (Shannon, 1948)指数进行计算,计算公式为:

式中,为种类数;P为第种鱼类占总生物量的比例。
1.2.5 多元统计分析 为减少稀有种对相似性分析的干扰,分析前,首先剔除出现率较低的种类,即将仅在1个站位出现的种类剔除(管伟等, 2017)。将鱼类生物量数据进行平方根转换,计算Bray-Curtis (Bray, 1957)相似性系数矩阵,采用等级聚类分析(CLUSTER)来研究舟山沿岸渔场鱼类群落格局。用胁迫系数(Stress)来衡量NMDS二维点阵图结果的好坏,通常认为0.1≤Stress<0.2时,可用NMDS的二维点图表示,其图形有一定的解释意义;0.05≤Stress<0.1时,排序效果基本可信;可以认为是一个好的排序;Stress<0.05时,具有很好的代表性(夏陆军等, 2016)。利用ANOSIM检验分析群落间的差异性。采用相似性百分比(SIMPER)分析组内相似性和组间相异性及典型种和分歧种的贡献率。
1.2.6 RDA分析 在CANOCO4.5软件中分析鱼类群落与环境因子的关系,来判断对鱼类群落影响最显著的环境因子。在确定使用何种方法分析鱼类群落和环境因子的关系前,先用species-sample数据进行DCA分析,如果所得结果中Lengths of gradient第一排序轴的值大于4.0,则选CCA分析(即典范对应分析);如果其值在3.0~4.0之间,则选CCA分析或RDA分析(即冗余分析)均可以;如果其值小于3.0,则RDA分析所得结果会优于CCA分析(Ter Braak, 1986、1988)。
调查站位图以及多样性指数平面分布图通过Surfer11绘制。等级聚类及标序分析、ANOSIM检验、相似性百分比分析均由PRIMER5.0完成。
2 结果
2.1 群落组成
春季和秋季共鉴定出鱼类81种,隶属于12目39科63属,其中,春季有58种,秋季有53种,分别占总种类数的71.60%和65.43%。按鱼类适温性及适盐性,调查海域鱼类分为近海暖水种、近海暖温种、沿岸暖水种、沿岸暖温种、河口暖温种、近海冷温种和河口暖水种7种生态类群,各生态类型总种数和在各季节的种类数及其所占百分比如表1所示。其中,以近海暖水种最多,共25种,占鱼类种数的30.86%,春、秋季分别为18和15种,分别占31.03%和28.30%;河口暖水种鱼类最少,仅2种,占鱼类种数的2.47%,春、秋季分别有2种和1种,各占3.45%和1.89%(表1)。
春、秋季共有种类30种,种类相似性指数为37.04%,显示2个季节的种类组成有较大变化,种类组成季节差异性较大。
2.2 优势种的季节变化
根据优势种计算结果可知,本次调查春季优势种为六丝钝尾虾虎鱼(Amblychaeturichthys hexanema),其重量占春季鱼类总重量的42.16%,尾数占总尾数的63.36%,优势度远高于其他种类,常见种有棘头梅童鱼(Collichthys lucidus)、褐菖鲉(Sebastiscus marmoratus)和蓝圆鲹(Decapterus maruadsi);秋季优势种为龙头鱼(Harpodon nehereus),占春季鱼类总重量的43.56%,占总尾数的80.52%,所有站位均捕获到该种类。本调查捕获的主要鱼类营养级介于3.4~4.2 (纪炜炜, 2011; http://www.fishbase.se/search.php.),总体表现为中级(表2)。
2.3 鱼类群落结构
对春季和秋季鱼类的群落结构进行等级聚类分析(CLUSTER)和NMDS分析(图2和图3),2个季节NMDS排序的胁迫系数分别为0.14和0.11,均在0.1~0.2之间,NMDS排序的二维点阵图结果是可以接受的。各季节站位聚类分析和NMDS分析结果基本一致,春季该海域的鱼类可划分为群落Ⅰ、Ⅱ、Ⅲ和Ⅳ4个组群;秋季该海域的鱼类可划分为群落Ⅰ、Ⅱ和Ⅲ3个组群。
表1 各生态类型种类数及所占百分比

Tab.1 Number and percentage of each ecological type
表2 舟山沿岸渔场主要鱼类生态位宽度及营养级

Tab.2 The niche width and trophic level of major fish in the surveyarea
注:“–”表示未查到的营养级
Note: “–”refers to the nutrition level not found

图2 鱼类群落Bray-Curtis相似性聚类

图3 鱼类群落结构的NMDS分析
如表3所示,春季群落Ⅰ中的典型种为蓝圆鲹、棘头梅童鱼和龙头鱼;群落Ⅱ中的典型种为棘头梅童鱼、褐菖鲉、中华小公鱼(Stolephorus chinensis);群落Ⅲ中的典型种为蓝圆鲹和六丝钝尾虾虎鱼;群落Ⅳ中的典型种为棘头梅童鱼和六丝钝尾虾虎鱼。如表5所示,秋季群落Ⅰ、Ⅱ中的典型种为龙头鱼;群落Ⅲ中的典型种为龙头鱼、中华小沙丁鱼(Sardinella nymphaea)、鳀()、江口小公鱼()和七星底灯鱼(Benthosema pterotum)。
如表4所示,春季,群落Ⅰ、Ⅱ的相异性为91.60%,主要分歧种为蓝圆鲹、棘头梅童鱼和龙头鱼等;群落Ⅰ、Ⅲ的相异性为97.04%,主要分歧种为蓝圆鲹、棘头梅童鱼和龙头鱼等;群落Ⅰ、Ⅳ的相异性为90.58%,主要分歧种为蓝圆鲹、棘头梅童鱼、六丝钝尾虾虎鱼和龙头鱼等;群落Ⅱ、Ⅲ的相异性为86.71%,主要分歧种为蓝圆鲹、棘头梅童鱼、六丝钝尾虾虎鱼、褐菖鲉、中华小公鱼和小带鱼等;群落Ⅱ、Ⅳ的相异性为82.62%,主要分歧种为棘头梅童鱼、六丝钝尾虾虎鱼、褐菖鲉和中华小公鱼等;群落Ⅲ、Ⅳ的相异性为80.88%,主要分歧种为棘头梅童鱼、六丝钝尾虾虎鱼和褐菖鲉等。如表6所示,秋季,群落Ⅰ、Ⅱ的相异性为81.75%,主要分歧种为龙头鱼、黄鲫(Setipnna taty)、红鳗虾虎鱼(Odontamblyopus rubicundus)、棘头梅童鱼、六丝钝尾虾虎鱼和中华小沙丁鱼等;群落Ⅰ、Ⅲ的相异性为93.46%,主要分歧种为龙头鱼、黄鲫、红鳗虾虎鱼、棘头梅童鱼、六丝钝尾虾虎鱼和七星底灯鱼等;群落Ⅱ和群落Ⅲ的相异性为74.55%,主要分歧种为龙头鱼、中华小沙丁鱼、七星底灯鱼、鳀、江口小公鱼等。
表3 春季鱼类群落组内典型种及贡献率(%)

Tab.3 Typifying species within station groups and their contributions percentage for fish community structure in spring (%)
注:Av.Sim表示平均相似性;Con表示贡献率。下同
Note: Av.Sim: Average similarity; Con: Rate of contribution. The same as below
表4 春季鱼类群落组间分歧种及贡献率(%)

Tab.4 Discriminating species between spatial groups and their contributions percentage for fish community structure in spring (%)
注:Av.Diss表示平均差异性。下同
Note: Av.Diss: Average dissimilarity. The same as below
表5 秋季鱼类群落组内典型种及贡献率(%)
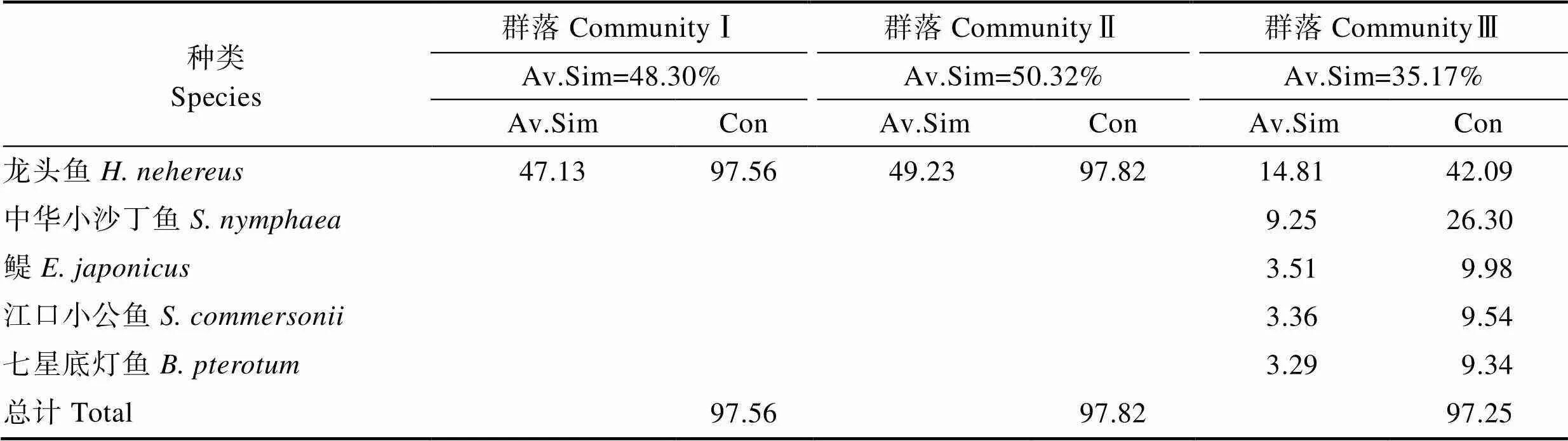
Tab.5 Typical species and their contribution rate among fish community in autumn(%)
表6 秋季鱼类群落组间分歧种及贡献率(%)

Tab.6 Different species and their contribution rate among fish community in autumn (%)
2.4 鱼类生物多样性
春季,鱼类多样性指数(′)为1.75±0.64,以13号站位最高,′值为3.13,以75号站位最低,′值为0.13;秋季,鱼类多样性指数′值为0.93±0.74,以4号站位最高,′值为2.94,以72号站位最低,′值为0.01 (图4)。
2.5 鱼类群落与环境因子的关系
对春、秋季species-sample数据进行DCA分析,结果显示,Lengths of gradient的第1排序轴的大小均小于3.0,因此,两季均采用RDA分析方法研究鱼类群落和环境因子的关系。
对春、秋季舟山沿岸渔场调查站位RDA排序,春、秋季鱼类群落与环境因子的RDA排序与NMDS的排序结果一致。其中,春季第1排序轴与第2排序轴的特征值分别为0.2730和0.0260,相关系数分别为0.7990和0.5240,解释了物种变异的28.60%和环境变异的86.15%。秋季第1排序轴与第2排序轴的特征值分别为0.1320和0.0240,相关系数为0.6330和0.5290,解释了物种变异的14.40%和环境变异的81.55%。说明春、秋季排序轴的相关性都很小,排序结果可靠(表7)。
春季,第1排序轴与溶解氧的正相关系数最高,与表层盐度呈现出最大负相关。由此可知,在排序图中,化学耗氧量(COD)从左往右逐渐升高,而表层水温、底层水温、表层盐度、底层盐度和叶绿素逐渐降低。叶绿素与第2排序轴的正相关系数最大,表层盐度与第2排序轴呈最大负相关。由此可知,叶绿素由下至上逐渐增加,表层盐度和表层水温逐渐减少(图5)。

图4 生物多样性空间分布
表7 舟山沿岸渔场春、秋季排序轴特征值以及其和环境变量的相关系数

Tab.7 The eigenvalues of axes and their correlation coefficients with environment variables in Zhoushan coastal fishing ground during two seasons

图5 调查海域鱼类站位与环境因子的RDA排序
秋季,表层盐度与第1排序轴呈最大负相关,叶绿素与第1排序轴呈最大正相关。由此可知,在排序图中,表层盐度、水深、表层水温、底层水温和底层盐度从左往右逐渐降低,叶绿素与COD逐渐增大。表层水温和第2排序轴表现为最大正相关,表层盐度和第2排序轴表现为最大负相关。由此可见,表层水温由下至上逐渐增加,表层盐度、底层盐度和叶绿素逐渐降低(图5)。
3 讨论
3.1 鱼类种类及优势种组成分析
本调查中,春季有58种鱼类,秋季有53种鱼类。将本调查结果与2006~2007年俞存根等(2010)的调查结果相比可发现,2006~2007年该海域春季有75种鱼类,秋季有79种鱼类,2次调查结果相差很大,其原因不仅与调查范围、调查海域的海洋环境有关,也与调查网具、过度捕捞造成渔业资源衰退等有关。其中,过度捕捞也是鱼类群落营养级降低的主要原因,随着鱼类群落演替的发生,低营养级逐渐取代高营养级,使可捕鱼类有小型化、低值化的趋势,如六丝钝尾虾虎鱼等(纪炜炜, 2011)。
舟山群岛位于北亚热带南缘,受东亚季风影响,该海域周围散布着上千个岛屿,水文环境非常独特(侯伟芬等, 2013),又受到杭州湾和长江口以及暖寒流的影响,使海水盐度出现较大的变化。因此,会按照不同盐度梯度出现不同的生态类型(王迎宾等, 2012)。从鱼类生态习性来看,本调查以近海种最多,河口种最少,暖水种最多,冷温种最少。可能是因为调查海域受长江径流、江浙沿岸流和台湾暖流的影响(张启龙等, 2004),导致该海域营养盐丰富,饵料生物充足,大量的近海种在此产卵和索饵;而冷温种都分布在调查海域北部,该海域受江浙沿岸流的影响显著,水温较低,适宜冷温种生存。春季和秋季又均以近海暖水种最多,并且以调查海域中部及南部海域站位捕获较多,主要原因是舟山沿岸渔场受到台湾暖流的影响,海水温度较高,给暖水种提供了适宜的生存环境。虽然大多数站位所处的海洋环境不同,但近海暖水种的种类基本相同,可见该海域鱼类种类多以广温广盐性为主。春季的优势种六丝钝尾虾虎鱼和秋季的优势种龙头鱼均属于广温广盐性的种类,这2种鱼类的生物量分别占生物总量的42.16%和43.56%,原因可能是整个调查海域海洋环境复杂,又受江浙沿岸流的影响显著(俞存根等, 2016),海水温度和盐度变化较大,但春季和秋季各优势种的出现率均在90%以上,秋季龙头鱼的出现率达到100%,可见这2种鱼类对环境的适应性较强。群聚相似性显示,2个季节的种类相似性并不高,主要原因可能是由于春季和秋季鱼类不同的洄游活动,以及鱼类对温度盐度的适应性,使2个季节捕获的鱼类种类存在差异。
3.2 鱼类多样性分析
从鱼类生物多样性的季节变化来看,春季生物多样性指数高于秋季,可能是由于春季是大多数鱼类的繁殖季节,亲鱼和幼鱼主要栖息于沿岸渔场,且近岸水域是多种渔业生物的重要产卵场与栖息地(俞存根等, 2016),因此,沿岸渔场生物多样性较高;而到了秋季,随着台湾暖流势力的减弱,导致该海域水温下降,致使一些鱼类会向外海深水区进行洄游,生物多样性降低,这与俞存根等(2010)研究的舟山渔场及邻近海域鱼类以洄游性种类或季节性种类为主的结论一致。2个季节的生物多样性指数呈相似的分布趋势(图5),均以调查海域北部最高,而以29.8°N以南范围最低,其次是杭州湾附近海域。岛屿附近多样性指数较高,尤其以嵊泗列岛和中街山列岛附近海域高于其他区域,其原因一方面可能是因为岛屿附近流态复杂,营养物质丰富,适宜多种鱼类生存,并且中街山列岛作为海洋保护区,该海域受到多方保护,从而维持了这一特殊海域的生态系统多样性(程郡等, 2014),另一方面也是因为岛屿之间特殊的海底地形、底质和水流环境多样化满足了不同鱼类的生活要求,使大多数鱼类有合适的栖息地(徐兆礼等, 2008),因此,岛屿附近的多样性较其他水域高。
3.3 鱼类群落结构与环境因子的关系
从等级聚类和多维标度排序分析结果可以看出,春季和秋季的二维标序图均有一定的解释意义。通常情况下,等级聚类和多维标度排序分析结果可以相互对照,这样得出的结论更具有生态学意义(厉红梅等, 2001)。舟山沿岸渔场春季可分为4个组群,秋季可分为3个组群,各组群典型种均以海洋性种类为主,且各组的典型种也是各季节的优势种或重要种,因此,其群落结构的空间差异主要是由于各群组种类组成不同及主要优势种数量的相对变化造成的(王小林等, 2013)。本研究表明,春季和秋季舟山沿岸渔场鱼类群落与底层水温和底层盐度均有显著相关性,这可能由舟山沿岸渔场复杂的地形特点与海流等因子的共同作用造成(唐逸民等, 1991),调查海域岛屿众多,底部的岩石容易使该海区产生上升流等,会影响该海域盐度等水文环境,并且该海域又受台湾暖流和黄海冷水团的影响,均会使该海域水温与盐度发生变化。根据RDA分析结果可知,调查海域温度、盐度、水深、叶绿素a等对鱼类群落结构有重要的影响。水深在海洋环境中是对鱼类群落结构变化有重要影响作用的环境因子(Colvocoresses, 1984),和鱼类群落结构的联系较为密切,不同深度海域具有不同的环境,环境因素的变化对鱼类分布有限制作用,不同环境中分布着不同的种类,从而形成鱼类群落(王雪辉等, 2010)。调查海域离大陆较近,且是江浙沿岸流与台湾暖流的交汇区(许建平等, 1992),是该海域水温、盐度因子的重要影响因素。春季底层盐度与叶绿素对鱼类群落的影响显著,这可能因为本次春季调查的时间在5月,正是雨水丰沛的时节,大量冲淡水会使海水中的盐度降低,而盐度又会影响鱼类的生理活动,如鱼类代谢及渗透压调节等,从而对鱼类群落结构造成影响。由于调查海域受径流的影响,径流会将大量来自陆源的物质,包括悬浮物以及溶解性营养盐带入海水中,从而使海水中浮游植物得以更好生长,因此,调查海域的叶绿素含量较高,浮游植物的大量生长又促进饵料生物的生长,间接影响了鱼类群落结构。从图5还可以看出,春季位于杭州湾附近海域调查站位的鱼类群落受盐度的影响明显,这与大量的沿岸冲淡水有关。秋季温度、盐度及叶绿素总体对调查海域鱼类群落结构的影响较大,主要与流经此处的高温高盐台湾暖流势力减弱、海区水温与盐度均有所降低有关。秋季调查海域偏北部及外部海域站位的鱼类群落受叶绿素的影响较大,外部海域的透明度较高,浮游植物光合作用充足,而岛屿附近调查站位的鱼类群落结构受调查海域温度、盐度和水深的影响较大,主要是因为岛屿附近受大陆径流的影响,盐度和温度均有较大的变化。水深也与盐度和水温的变化有相关性,可能是因为水深变化会使有伴随关系的底层温度和底层盐度发生变化(王雪辉等, 2010; 邱永松等, 1988; 杨涛等, 2018; 张波等, 2013)。
Bray JR, Curtis JT. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 1957, 27(4): 325–349
Cao DY, Yu CG, Zhou QS,. The species composition and distribution characteristics of fish in the waters nearby Putuo Mountain of Zhoushan fishing ground. Fujian Fisheries, 2015,37(5): 353–362 [曹东阳, 俞存根, 周青松, 等. 舟山渔场普陀山附近海域鱼类种类组成和数量分布. 福建水产, 2015, 37(5): 353–362]
Chen DG, Zhang MZ. Marine fish of China. Qingdao: China Ocean University Press, 2015, 12 [陈大刚, 张美昭. 中国海洋鱼类. 青岛: 中国海洋大学出版社, 2015, 12]
Chen JJ, Xu ZL, Wang CH. Fish community in Jintang Island sea area of Zhoushan fishing grounds, Zhejiang Province of East China in spring and summer. Chinese Journal of Ecology, 2012, 31(4): 931–938 [陈佳杰, 徐兆礼, 王翠华. 舟山渔场金塘岛海域春夏季的鱼类群落. 生态学杂志, 2012, 31(4): 931–938]
Cheng J, Zang YL, Yan LN. The restoration of fisheries resources in Zhoushan coastal water. Journal of Anhui Agricultural Sciences, 2014, 42(16): 5166–5167 [程郡, 臧迎亮, 闫丽娜. 舟山沿岸渔业资源修复. 安徽农业科学, 2014,42 (16): 5166–5167]
Colvocoresses JA, Musick JA. Species associations and community composition of middle Atlantic Bight continental shelf demersal fishes. Fish Bull, 1984, 82(2): 295−313
Froese R, Paulyd. Fish Base. http://www.fishbase.se/search. php
Guan W, Xu ZL, Chen JJ. Structure and diversity of fish communities in the waters south of Nanri Island. Acta Ecologica Sinica, 2017, 37(9): 3172−3181 [管伟, 徐兆礼, 陈佳杰. 福建南日岛南部水域鱼类群落结构及多样性. 生态学报, 2017, 37(9): 3172−3181
Hou WF, Yu CG, Chen XQ. Temperature distribution in Zhoushan fishing ground. Journal of Ningbo University(NS EE), 2013, 26(3): 31–34 [侯伟芬, 俞成根, 陈小庆. 舟山渔场的水温分布特征分析. 宁波大学学报(理工版), 2013, 26(3): 31– 34]
Ji WW. Ecological studies on the food web structures and trophic relationships of Northern and Central East China Sea using stable carbon and nitrogen isotopes. Doctoral Dissertation of Institute of Oceanology, Chinese Academy of Sciences, 2011 [纪炜炜.东海中北部主要游泳动物食物网结构和营养关系初步研究.中国科学院海洋研究所博士研究生学位论文, 2011]
Li HM, Cai LZ, Lin LZ,. Using hierarchial clustering and no-metric MDS to explore spatio-temporal variation of benthic community at intertidal in Shenzhen Bay. Journal of Xiamen University(Natural Science) 2001, 40(3): 735– 740 [厉红梅, 蔡立哲, 林丽珠, 等. 深圳湾潮间带底栖动物群落结构的等级聚类与非度量多维标度排序. 厦门大学学报(自然科学版), 2001, 40(3): 735–740]
Li J, Li XH, Jia XP,. Evolvement and diversity of fish community in Xijiang River. Journal of Fishery Sciences of China, 2010, 17(2): 298–311 [李捷, 李新辉, 贾晓平, 等. 西江鱼类群落多样性及其演变. 中国水产科学, 2010, 17(2): 298–311]
Li SF, Cheng JH, Li CS,. Seasonal changes on fish community diversity in the middle part of the East China Sea. Marine Fisheries, 2005, 27(2): 113–119 [李圣法, 程家骅, 李长松, 等. 东海中部鱼类群落多样性的季节变化. 海洋渔业, 2005, 27(2): 113–119]
Li SF, Cheng JH, Yan LP. Spatial structure of fish communities on the continental shelf of the East China Sea. Acta Ecologica Sinica, 2007, 27(11): 4377–4386 [李圣法, 程家骅, 严利平. 东海大陆架鱼类群落的空间结构. 生态学报, 2007, 27(11): 4377–4386]
Liang J, Wang WD, Yu BC,. Seasonal variation of fish resources and community diversity in reef habitat in marine protected area of Zhongjieshan islands. Oceanologia et Limnologia Sinica, 2014, 45(5): 979–989 [梁君, 王伟定, 虞宝存, 等. 中街山列岛海洋保护区岛礁生境鱼类资源及群落多样性季节变化. 海洋与湖沼, 2014, 45(5): 979–989]
Lin N, Miao ZQ, Lu ZH. Structure and diversity of fish communities in summer in the middle of the East China Sea. Journal of Zhanjiang Ocean University, 2009, 29(3): 42–47 [林楠, 苗振清, 卢占晖. 东海中部夏季鱼类群落结构及其多样性分析. 广东海洋大学学报, 2009, 29(3): 42– 47]
Lu ZH, Miao ZQ, Lin N. The structure and diversity fish communities in spring in the middle sea area of Zhejiang Province and adjacent region. Journal of Zhejiang Ocean University (Natural Science), 2009, 28(1): 51–56 [卢占晖, 苗振清, 林楠. 浙江中部近海及其邻近海域春季鱼类群落结构及其多样性. 浙江海洋学院学报(自然科学版), 2009, 28 (1): 51–56]
Qiu YS. The regional changes of fish communities on the northern continental shelf of South China Sea. Journal of Fisheries of China, 1988, 12(4): 303–313 [邱永松. 南海北部大陆架鱼类群落的区域性变化. 水产学报, 1988, 12(4): 303–313]
Quetglas A, Carbonell A, Sάnchez P. Demersal continental shelf and upper slope cephalopod assemblages from the Balearic Sea (North-Western Mediterranean). Biological aspects of some deep-sea species. Estuarine, Coastal and Shelf Science, 2000, 50(6): 739–749
Shan XJ, Chen YL, Dai FQ,. Variations in fish community structure and diversity in the sections of the central and southern Yellow Sea. Acta Ecologica Sinica, 2014, 34 (2): 377–389 [单秀娟, 陈云龙, 戴芳群, 等. 黄海中南部不同断面鱼类群落结构及其多样性. 生态学报, 2014, 34(2): 377–389]
Shannon EC, Weaver W. The mathematical theory of communication. Illinois: Urbana University of Illinois Press, 1948: 125
Shen JZ, Cheng YH. A study on the deep sea demersal fish communities and their structure in the East China Sea. Journal of Fisheries of China, 1987, 11(4): 293–306 [沈金鏊, 程炎宏. 东海深海底层鱼类群落及其结构的研究. 水产学报, 1987, 11(4): 293–306]
Sun SS, Tang WQ, Guo HY,Composition and changes in abundance and biomass of fish assemblages along the Jingjiang section of the Yangtze River over the last decade. Biodiversity Science, 2013, 21(6): 688–698 [孙莎莎, 唐文乔, 郭弘艺, 等. 靖江沿岸秋季鱼类群聚的组成特点及其丰度生物量变化. 生物多样性, 2013, 21(6): 688–698]
Tang YM. Oceanography. Beijing: China Agricultural Press, 1991 [唐逸民. 海洋学. 北京: 中国农业出版社, 1991]
Ter Braak CJF, Prentice IC. A theory of gradient analysis. Advances in Ecological Research, 1988, 18: 271–317
Ter Braak CJF. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167–1179
Wang XH, Qiu YS, Du FY,. Fish community pattern and its relation to environmental factors in Beibu Gulf. Journal of Fisheries of China, 2010, 34(10): 1579–1586 [王雪辉, 邱永松, 杜飞雁, 等. 北部湾鱼类群落格局及其与环境因子的关系. 水产学报, 2010, 34(10): 1579–1586]
Wang XL, Xu BD, Ji YP,. Fish community structure and its relationship with environmental factors in Haizhou Bay and adjacent waters of East China in winter . Journal of Applied Ecology, 2013, 24(6): 1707–1714 [王小林, 徐宾铎, 纪毓鹏, 等. 海州湾及邻近海域冬季鱼类群落结构及其与环境因子的关系. 应用生态学报, 2013, 24(6): 1707–1714]
Wang YB, Yu CG, Chen QZ,. Community structure of fish in Zhoushan fishing ground and its adjacent waters in spring and summer. Chinese Journal of Applied Ecology, 2012, 23(2): 545–551 [王迎宾, 俞存根, 陈全震, 等. 春、夏季舟山渔场及其邻近海域鱼类群落格局. 应用生态学报, 2012, 23(2): 545–551]
Wang ZH, Wang K, Zhao J,. Fish community structure and its seasonal change in subtidal sandy beach habitat off southern Gouqi Island. Chinese Journal of Applied Ecology, 2011, 22(5): 1332–1342 [汪振华, 王凯, 赵静, 等. 枸杞岛潮下带沙地生境鱼类群落结构和季节变化. 应用生态学报, 2011, 22(5): 1332–1342]
Xia LJ, Yu CG, Cai HC,. Community structure and diversity of shrimp in Nanji Islands marine conservation area. Acta Oceanologica Sinica, 2016, 38(2): 73–83 [夏陆军, 俞存根, 蔡厚才, 等. 南麂列岛海洋自然保护区虾类群落结构及其多样性. 海洋学报, 2016, 38(2): 73–83]
Xu JP, Yang SY. Hydrological and climatic characteristics of Nanji Islands and their adjacent waters. In: Nanji Islands National Marine Nature Reserve (1). Beijing: Marine Press, 1992, 1–9 [许建平, 杨士英. 南麂列岛及其附近海域的水文和气候特征. 南麂列岛国家海洋自然保护区论文选(一). 北京: 海洋出版社, 1992, 1–9]
Xu ZL. Analysis of fish diversity in the waters off the Oujiang estuary in summer and autumn. Acta Ecologica Sinica, 2008, 28(12): 5948–5956 [徐兆礼. 瓯江口海域夏秋季鱼类多样性. 生态学报, 2008, 28(12): 5948–5956]
Xu ZL. Comparison of fish fauna of two different waters (Minjiang Estuary and Xinghua Bay) of the East China Sea during spring and summer. Biodiversity Science, 2011, 19(1): 79–84 [徐兆礼. 兴化湾和闽江口海域春夏季鱼类区系特征. 生物多样性, 2011, 19(1): 79–84]
Yang T, Shan XJ, Jin XS,. Long-term changes in keystone species in fish community in spring in Laizhou Bay. Progress in Fishery Sciences, 2018, 39(1): 1–11 [杨涛, 单秀娟, 金显仕, 等. 莱州湾春季鱼类群落关键种的长期变化. 渔业科学进展, 2018, 39(1): 1–11]
Yu CG, Chen QZ, Chen XQ,. Species composition and quantitative distribution of fish in Zhoushan Fishing Ground and its adjacent waters. Oceanologia et Limnologia Sinica, 2010, 41(3): 410–417 [俞存根, 陈全震, 陈小庆, 等. 舟山渔场及邻近海域鱼类种类组成和数量分布. 海洋与湖沼, 2010, 41(3): 410–417]
Yu CG, Ye ZJ, Han ZQ. Fishery resources and fisheries science. Beijing: China Agricultural Press, 2016, 12 [俞存根, 叶振江, 韩志强. 渔业资源与渔场学. 北京: 中国农业出版社, 2016, 12]
Zhang B, Wu Q, Jin XS. Feeding ecology of fish assemblages and its variations in the Laizhou Bay. Progress in Fishery Sciences, 2013, 34(2): 1–9 [张波, 吴强, 金显仕. 莱州湾鱼类群落的营养结构及其变化. 渔业科学进展, 2013, 34(2): 1–9]
Zhang HL, Xu DK, Zhu ZJ,. Seasonal variations of fish resources and its community diversity in Daiquyang. Journal of Fisheriesof China, 2012, 36(4): 601–607 [张洪亮, 徐开达, 朱增军, 等. 岱衢洋鱼类资源及其群落多样性的季节变化. 水产学报, 2012, 36(4): 601–607]
Zhang QL, Wang F. Climatological analysis of water masses in Zhoushan fishing ground and adjacent region. Oceanologia et Limnologia Sinica, 2004, 35(1): 48–54 [张启龙, 王凡. 舟山渔场及其邻近海域水团的气候学分析. 海洋与湖沼, 2004, 35(1): 48–54]
Zhong YF Yu CG, Zhou QS,. Species composition and quantity distribution of fish in Ningbo-Zhoushan port. Journal of Zhejiang Ocean University (Natural Science), 2016, 35(1): 1–7 [钟毅飞, 俞存根, 周青松, 等. 舟山渔场宁波–舟山港海域春秋季鱼类种类组成和数量分布. 浙江海洋学院学报(自然科学版), 2016, 35(1): 1–7]
Analysis of the Fish Community Structure in the Spring and Autumn of the Zhoushan Coastal Fishing Grounds
GUO Pengjun1,2, JIANG Xinqin2, YU Cungen2①, DENG Xiaoyan2, ZHANG Ping2, XU Yongjiu2
(1. Zhoushan Marine and Fisheries Bureau, Zhoushan 316000; 2. Zhejiang Ocean University, Zhoushan 316022)
Species composition, ecological role, diversity, and clustering characteristics of fish species were analyzed by means of the species similarity, diversity index, and redundancy analysis (RDA) fishery resource surveys data collected in November 2015 (autumn) and May 2016 (spring) in the Zhoushan coastal fishing ground. The results showed that a total of 81 fish species were identified, which belonged to 63 genera in 39 families of 12 orders, with the bulk belonging to offshore, warm-water species. The Jaccard similarity index is 37.04% at the middle dissimilarity level. Analysis showed that the dominant species in the spring was Amblychaeturichthys hexanema, which accounted for 42.16% of the total catches. The dominant species in the autumn was Harpodon nehereus, which accounted for 43.56% of the total catches. The diversity index value of spring was higher than in autumn. However, the two seasonal diversity indices had similar planar distribution. Diversity was observed to be the lowest in the southern region of the survey area increasing toward the north, and a similar trend was seen from east to west in the survey region. The analysis of similarities (ANOSIM) test showed that the differences between the communities were highly significant. The similarity percentage (SIMPER) showed that seven species of fish, including Decapterus maruadsi, Collichthys lucidus,and H. nehereus, were typical species belonging to different groups in the spring. Fish community structure differed in autumn compared to spring primarily in that there were nine species of fish, including Sardinella nymphaea, Engraulis japonicus, and Benthosema pterotum. The results of the RDA indicated that the main environmental factors influencing the structure of fish community in the Zhoushan coastal fishing grounds were salinity and temperature.
Fish; Species composition; Diversity; Community structure; Zhoushan coastal fishing grounds
YU Cungen, E-mail: cgyu@zjou.edu.cn
S932
A
2095-9869(2020)04-0001-11
10.19663/j.issn2095-9869.20190311001
http://www.yykxjz.cn/
郭朋军, 江新琴, 俞存根, 邓小艳, 张平, 许永久. 舟山沿岸渔场春秋季鱼类群落结构特征分析. 渔业科学进展, 2020, 41(4): 01–11
Guo PJ, Jiang XQ, Yu CG, Deng XY, Zhang P, Xu YJ. Analysis of the fish community structure in the spring and autumn of the Zhoushan coastal fishing grounds. Progress in Fishery Sciences, 2020, 41(4): 01–11
* 国家自然科学基金项目(31270527)和舟山市海洋与渔业局委托项目(2015–2017年,海洋工程项目建设对舟山近岸海域渔业生态损害的补偿等级划分)共同资助 [This work was supported by National Natural Science Foundation (31270527), and Project Commissioned by Zhoushan Marine and Fisheries Bureau (Classification of Compensation Level for Fishery Ecological Damage in Zhoushan Coastal Area by Marine Engineering Project Construction in 2015–2017)]. 郭朋军,E-mail: 93658486@qq.com
俞存根,教授,E-mail: cgyu@zjou.edu.cn
2019-03-11,
2019-05-28
(编辑 马璀艳)
猜你喜欢
中国水产(2021年12期)2021-12-06
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
大自然探索(2021年12期)2021-02-07
渔业研究(2020年4期)2020-08-21
河南水产(2020年1期)2020-06-29
上海农业学报(2020年2期)2020-05-04
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
金山(2018年8期)2018-11-09
