
傅里叶变换红外显微光谱(Micro-FTIR)和X 射线衍射(XRD)用于测定棉花结晶度效果比较
2020-07-23 03:15张李鹏张石定许鹏李现常张震范森淼龚举武袁有禄商海红邹华文
棉花学报 2020年4期
张李鹏,张石定,许鹏,李现常*,张震,范森淼,龚举武,袁有禄,商海红*,邹华文
(1.长江大学,湖北 荆州434025;2. 中国农业科学院棉花研究所,河南 安阳455000;3. 安阳工学院,河南 安阳455000)
纤维素结晶度(Crystalline index,CI)用于描述纤维素中结晶成分的含量,可以表征纤维内链状高分子有序排列的程度。 因此,CI 在一定程度上反映了纤维的物理力学、化学结构信息,是评价纤维材料性能和品质的重要依据[1-4]。 棉花作为重要的天然纤维材料,其成熟纤维中纤维素含量超过90%[5], 其纤维细胞的发育大致可以分为4个有重叠的阶段:纤维起始、初生壁形成(伸长)、次生壁加厚(纤维素的生物合成)以及成熟[6-8]。 研究棉花纤维在整个发育过程中结晶度的变化有助于得到其从无定形的凝胶状态到有序纤维的动态信息,该信息对于棉花的育种和纺织都有很重要的意义。
对于纤维素结晶度的检测,公认比较直接和可信的方法是使用X 射线衍射 (X-ray diffraction,XRD)[8-9]。 XRD 测量植物纤维结晶度的研究已经有很多年[10]。 使用XRD 研究纤维结晶度,主要有3 种方法[9]。方法一是使用XRD 衍射图中结晶成分峰强度(I200-Iam)与总强度(I200)的比值。 这种方法计算简单,可以快速计算纤维材料的结晶度,但由于峰强中包含非晶成分,很容易高估材料的结晶度。 方法二是使用总衍射强度扣除非晶衍射强度,以此获取结晶材料的衍射强度,最终计算出结晶度。 这个方法需要选择标准非晶样品,很难确定所选标准非晶样品同测试样品有相似的非结晶含量。 方法三是使用曲线拟合的方法, 把衍射图中的结晶成分与非晶成分拟合出来,然后计算结晶度。 由于样品成分复杂,衍射峰重叠区域广,拟合难度较大。虽然XRD 用于研究结晶材料效果显著,但并不能显示出分子结构的信息,如纤维内部的氢键作用信息等,而且结晶度值会由于方法不同而有差异。 因而,从另外一种技术角度寻找和建立新的更简洁、 更快速、更准确的CI 测量方法, 对促进和加深对纤维素结晶度的探索与研究具有重要意义。
傅里叶变换红外光谱 (Fourier transform infrared spectroscopy,FTIR)为分子振动光谱技术,可以获得样本的分子振动信息,并且光谱强度与样本中所含成分的含量成正比。 这项技术由于其快速、无损以及需样品量少等优点,已被越来越广泛地用于生物材料分析[8,10-15]。 近些年来,傅里叶变换红外光谱分析已经成为研究棉花纤维的常用手段[8,13-15],并被认为是计算纤维素结晶度最简单的方法[9]。 Devashankar 使用红外与XRD 和热重分析(Thermogravimetric analysis,TGA)联用测试木材样品,对木材的热动力学等性质有更深的理解[10]。而Abidi 等使用XRD 与红外联用的方法研究了不同时期棉花的发育状态, 将XRD 结晶度与红外中1 620 cm-1的峰强度相关联,得到红外峰强与XRD 结晶度之间的关系[8]。
利用红外光谱解析纤维素结晶度一般使用其相对峰强比值来计算。 以前报道中使用FTIR计算纤维素结晶度的方法包括Total Crystalline Index(FTIR-TCI)、Lateral Order Index(FTIR-LOI)、Hydrogen-Bond Intensity(FTIR-HBI)、Carrillo-Colom Index(FTIR-CCI)等[1-4,6,13]。 FTIR-TCI 和FTIR-LOI, 分别用α1376/α2902 和α1420/α893计算[1-2];FTIR-HBI,用α3336/α1336 计算[3];FTIRCCI,用α1278/α1263 计算[4]。 上述4 种方法是基于纯度较高的纤维素样本建立起来的方法,能否适用于棉花纤维发育过程还有待进一步研究。
本研究选取陆地棉(0-153)和海岛棉(S-6)作为研究对象, 分别取2 个材料开花后5、10、15、20、25 和30 d 的棉花纤维。 常规FTIR 方法可以获得样本宏观成分信息,而采用傅里叶变换红外显微光谱(micro-FTIR)则可以获得棉花纤维更深层次微观信息。鉴于此,采用micro-FTIR 与XRD联用技术,研究2 种棉花在不同时期的结晶度变化,并把FTIR-CI 与XRD-CI 作回归拟合相关性分析,验证FTIR-CI 与XRD-CI 拟合建立的显微红外光谱结晶度模型是否适用于成熟棉花纤维的结晶度计算。
1 材料与方法
1.1 材料
分别选取陆地棉0-153 和海岛棉S-6, 种植于中国农业科学院棉花研究所安阳实验基地(河南省安阳县白壁镇)。 分别取所选2 个棉花品系开花后5、10、15、20、25 和30 d 的棉铃, 使用手术刀剥去外皮,用镊子夹取纤维,处理发育前期的纤维时需要注意避免混入胚珠。 将剥好的纤维样品放入写好编号的2 mL 离心管中, 离心管于-80 ℃冰箱中保存,留待取用。
1.2 研究方法
1.2.1 材料处理。 将棉花纤维充分解冻后进行清洗。 在离心管中注入1.5 mL 超纯水,充分超声振荡,使纤维样品中可溶性糖等充分溶解,静置,倒掉上清液。 清洗操作重复3 次。 对于发育前期纤维样品,由于呈现胶状,清洗时会悬浮于超纯水中,倒上清液时易漏出,可以适当离心使纤维样品凝集于离心管底部。 清洗完成后,将棉花纤维慢慢从离心管底部移到靠近管口位置,开盖平放入40 ℃烘箱中48 h, 然后放入少量无水硫酸铜保持干燥,放置于温度为(21±1)℃环境内保存。每个离心管的样品代表一个品系的一个时期,并平均分为3 份,每份都随机选取纤维进行测试。1.2.2 傅里叶变换红外显微光谱。 红外光谱由日本岛津IRTracer-100 光谱仪采集,同时配备有液氮冷却MCT 探测器的岛津AIM 9000 显微镜。每个发育时期的纤维样品都分别固定在中间带有金刚石透明薄片的金属板上进行红外光谱采集。采用15 倍透镜,透射模式,波数范围为4 000~700 cm-1,分辨率为4 cm-1。每个品系的每个时期样本打乱铺平后随机选取3 根棉纤维(开花后5 d、10 d 样本为薄片状,随机选择3 片),每根棉纤维样本再选取3 个不同点采样,每点20 次扫描。 测试后使用大气CO2的光谱对所有红外光谱进行校正,并取平均光谱为每一品系在一个时期的最终光谱。
1.2.3 XRD 光谱。 XRD 数据是用帕纳科Empyrean X 射线衍射仪获得。Cu Kα 在加速电压为40 kV、电流为40 mA 条件下产生X 射线。 样品放置于中间有正方形凹槽的玻璃片上,凹槽大小约2 cm×2 cm。 鉴于发育前期的纤维样品,纤维形态呈现为胶状,趋向于相互黏连,干燥脱水后成薄片状, 样品可直接放置于玻璃片凹槽上。对于发育后期的纤维样品,可以将纤维平行密集排列固定到玻璃片上,在玻璃片中间凹槽手动压片。扫描2θ 角范围为10°~50°,步长为0.013 13°。
1.3 数据整理
1.3.1 红外数据。 利用Labsolutions 光谱软件多点自动基线校正方法对光谱进行平滑和基线扣除操作。 对于光谱平滑,所选点越多平滑程度越明显,但是平滑后的效果就越失真,所以需要选择合适点数对红外光谱进行平滑。 基线扣除完成后,导出每个棉花材料不同时期不同位置所获得的红外光谱原始ASCII 文件和扣完基线的ASCII 文件。
将ASCII 文件导入Microsoft Excel 2016 中,开始数值计算。 使用公式α=-lgT 将透射值转换为吸收值, 其中α 是吸收值,T 代表透射百分比(%)。 数值转换后,用MATLAB R2018a 对各个时期的光谱数据进行面积归一化,再求得各发育时期的平均值。 找出计算结晶度所需要的吸收峰 值,根 据 文 献[1-4]所 述FTIR-TCI、FTIR-LOI、FTIR-HBI、FTIR-CCI 方法分别计算结晶度。 将需要绘图数据导入Origin 9.0 进行绘图。
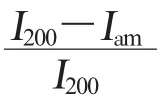
2 结果与分析
2.1 FTIR 结果
图1(a)是海岛棉S-6 在开花后25 d 白光成像图及测试选点, 其对应的透射模式FTIR 光谱如图1(b)。图2 所列是陆地棉0-153(a)和海岛棉S-6(b)随发育时期变化的FTIR 光谱图,部分有变化的红外峰已标示。 根据文献[7,14,17-18],红外峰的指认和归属见表1。
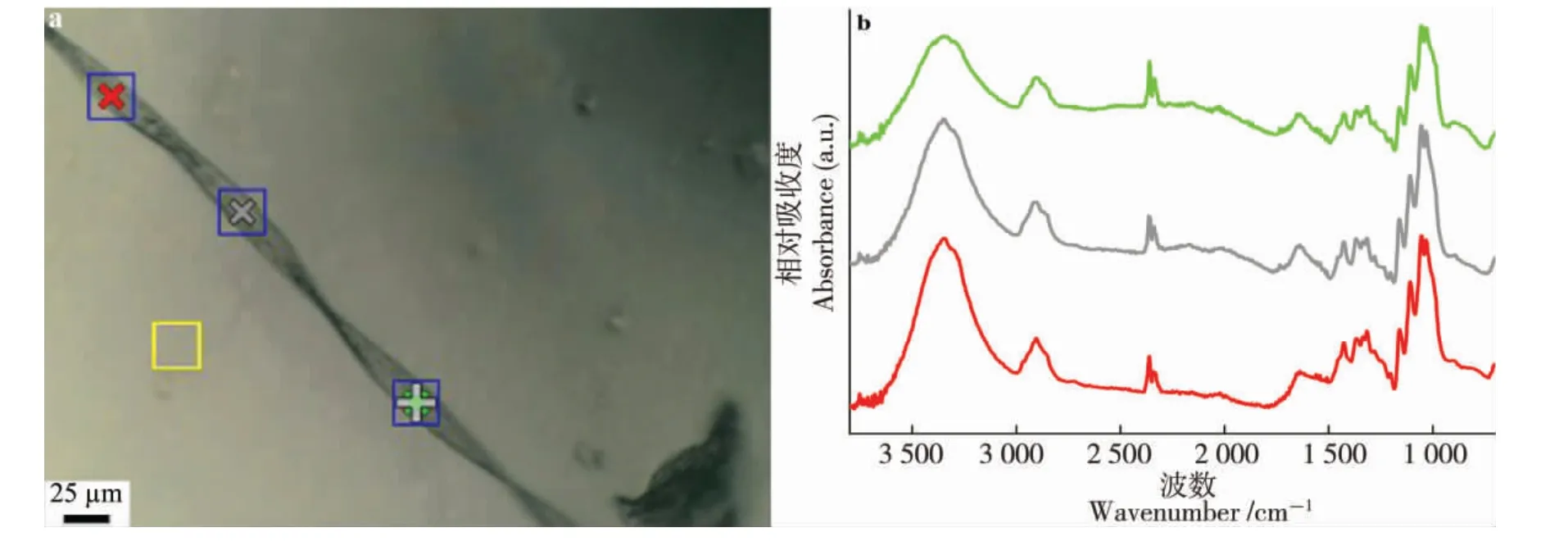
图1 白光图及其对应光谱图Fig. 1 Sampling points image and their spectra
在1 800~700 cm-1红外光谱区域, 有3 个吸收峰的相对强度变化值得注意, 位置分别在1 745、1 640 和1 543 cm-1,其强度随着棉花纤维发育而呈现逐渐降低甚至消失的趋势(图2)。1 640 cm-1附近的峰在开花后15 d 以后向1 620 cm-1偏移。 1 745 和1 640 cm-1峰归属于碳氧双键振动,来源于酯类、蛋白质或果胶物质。 上述峰的变化趋势表明,果胶等物质含量在棉纤维发育过程中是快速降低的,尤其在开花后15 d 以后。同时,1 242 cm-1附近的峰也来源于果胶类物质, 在开花后15 d 以后,峰强骤然变弱。 在1 543 cm-1附近的峰,被指认为NH2变形振动,来源于蛋白质或氨基酸。 1 543 cm-1在开花后5~15 d 持续降低,在开花后20 d 后几乎消失。 Abidi 等的研究有相同的现象[14],在开花后36 d 后出现检测不到的情况,但与本研究出现时期不同。 造成这种差异的原因可能是所选取的棉花材料或栽培环境不同。
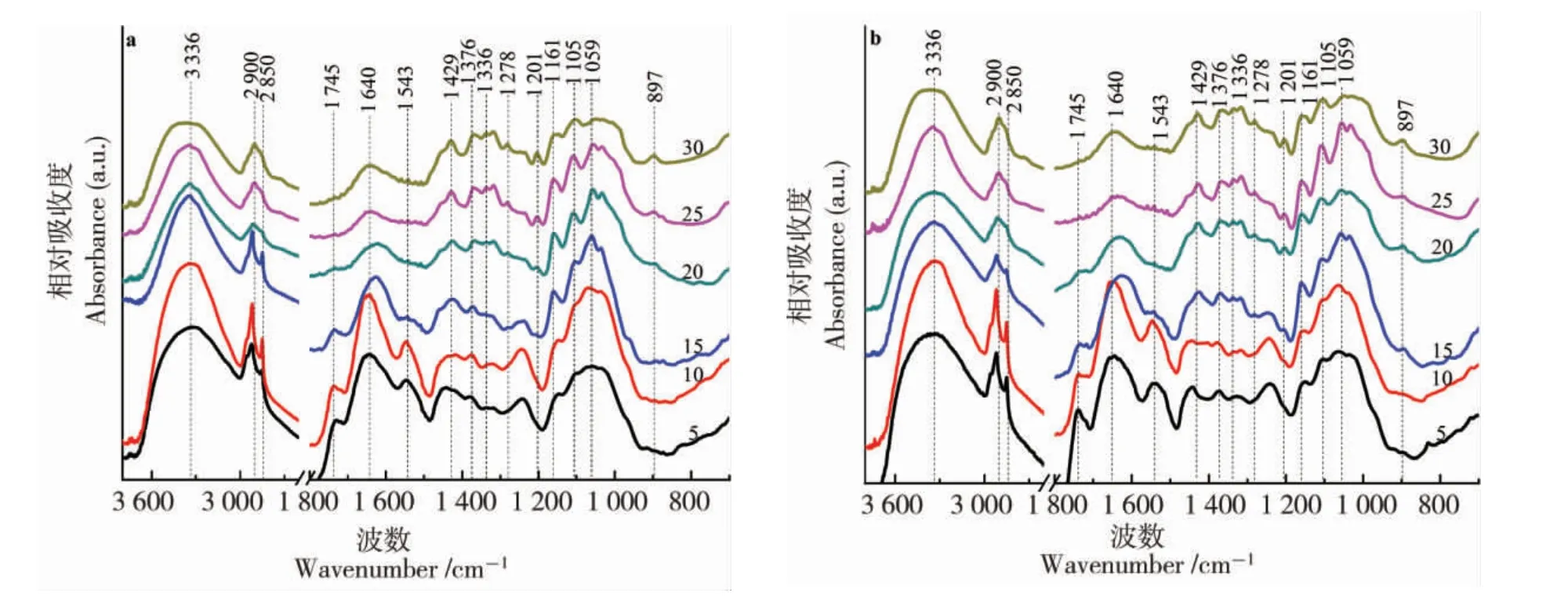
图2 0-153 和S-6 不同时期FTIR 光谱图Fig. 2 FTIR spectra in different developmental stages of 0-153 and S-6

表1 红外谱峰指认和来源Table 1 Selected peak assignments and origination
1 376 cm-1代表C-H 弯曲振动。 与1 640 cm-1相比,1 376 cm-1在开花后15 d 后逐渐增强。 1 315 cm-1代表CH2摆动,也有类似趋势。1 161 cm-1,代表C-O-C 的反对称拉伸模式,来源于纤维素。 1 105 cm-1代表平面内的反对称拉伸振动。 1 059 cm-1是C-O 的拉伸模式,在开花后15 d 后向低波数区域偏移。 1 240 cm-1吸收峰来源于果胶,在开花后15 d 前峰强较大,开花后20 d 后逐渐减小或消失。
1 429、1 336、1 278、1 201 和897 cm-1峰在开花后5、10 d 非常弱,后期逐渐变强(图2)。 对于1 429 cm-1峰, 在开花后15、20 d 处于1 425 cm-1位置,后期逐渐向1 429 cm-1偏移。 这些峰的逐渐增强可能与纤维素持续累积、含量增多密切相关。 在开花后15 d 时, 海岛棉S-6 的897 cm-1相对峰强度比陆地棉0-153 更高,反映出海岛棉S-6 棉纤维次生壁纤维素合成更多,可以推断在开花后15 d 之前海岛棉S-6 比陆地棉0-153的纤维发育快。
2.2 FTIR-CI 结果
图3 是2 个材料棉花纤维4 种方法得到的FTIR-CI 随棉纤维发育变化曲线。图3a 所示为陆地棉0-153 和海岛棉S-6 的FTIR-HBI 值, 峰强比值为α3336/α1336。 3 336 cm-1归属于纤维素的分子内部氢键的拉伸振动,1 336 cm-1被指认为O-H 平面内变形振动。该比值与纤维素中有序结晶相和分子排列的规律程度有关。 通常,FTIRHBI 值会随结晶度的上升而下降。 如图3a 所示,2 个材料的FTIR-HBI 值都随开花后时间增加而下降,在开花后30 d 后2 个材料降到了1.1。

图3 0-153 和S-6 的4 种FTIR-CI 随开花后时间的变化曲线Fig. 3 Change of FTIR-CI curve for 0-153 and S-6 fibers with the time after anthesis
图3b 所示为FTIR-LOI (1 429 与897 cm-1的2 个峰强比值) 随开花后时间的变化趋势。1 429 cm-1代表CH2对称弯曲振动,897 cm-1代表纤维素Iβ的C-O-C 拉伸振动,这2 个峰与纤维素含量变化密切相关。 该计算方法由O’Connor提出[1],随后被Hurtubise 和Krssig 称作Lateral Order Index(FTIR-LOI)[19]。 通常高的结晶度会有高的FTIR-LOI 值, 结晶度应该随着棉花纤维发育进程而增加。 但如图3b 所示,2 个材料FTIR-LOI 值都出现下降趋势。对于结晶度增加而FTIR-LOI 值下降的情况已有文献[4]提到。 这种现象的原因可能是发育阶段的棉花纤维由多种成分组成,FTIR-LOI 方法则是建立在纯度较高的结晶纤维素上的,可能不适合研究混合物样本[20]。
图3c 所示为FTIR-TCI 值 (1 376 与2 900 cm-1的2 个峰强比值) 随棉纤维发育的变化曲线。 1 376 cm-1属于纤维素的C-H 变形振动,2 900 cm-1属于C-H 拉伸振动模式,在结晶纤维素与非晶纤维素中都存在。 这个比值可以用来表征纤维素类物质结晶成分的比例。 从图3c 可以看出,2 个品系的FTIR-TCI 值都是随着开花后时间的增加而变大。
图3d 所示为FTIR-CCI 值 (1 278 与1 263 cm-1的2 个峰强比值) 随纤维发育的变化曲线。1 278 cm-1属于纤维素的C-H 变形振动,1 263 cm-1属于C-O-H 弯曲振动。 2 个品系棉纤维的FTIR-CCI 值随开花后时间的增加都表现出上升的趋势, 海岛棉S-6 在开花后10~15 d 时增长快,陆地棉0-153 则在开花后15~20 d 增长较快。
2.3 XRD 结果
从图4 可以明显看到,在10°~50°之间,有4个衍射峰的强度随纤维发育不断增强。 4 个衍射峰代表4 个晶面,分别是(110)晶面、(110)晶面、(200)晶面和(004)晶面,其中(200)晶面衍射强度最强,(004)晶面强度较弱。 所选的2 个棉花品系棉纤维的衍射峰具有相似的趋势: 开花后5 d时都只有(200)晶面衍射强度,开花后30 d 时都有4 个晶面衍射强度,但是衍射峰出现的时期和强度在2 个品系间有差异。 开花后15 d 时0-153只有(110)、(110)和(200)晶面强度,但S-6 已经出现(004)晶面衍射强度。 在开花后20 d,与陆地棉0-153 的各晶面衍射强度相比, 海岛棉S-6 的明显较弱,可能是由于海岛棉S-6 在此时及其以后较陆地棉0-153 发育慢。

图4 0-153 和S-6 不同纤维发育时期XRD-CIFig. 4 XRD-CI of 0-153 and S-6 fibers in different developmental periods
2.4 XRD-CI 结果
由图5 可以看出,在开花后5~15 d,海岛棉S-6 和陆地棉0-153 的XRD-CI 值非常相近,但在开花后20 d 时0-153 的XRD-CI 值稍微有所上升。开花后25 d 后,陆地棉0-153 与海岛棉S-6的XRD-CI 值又非常接近。
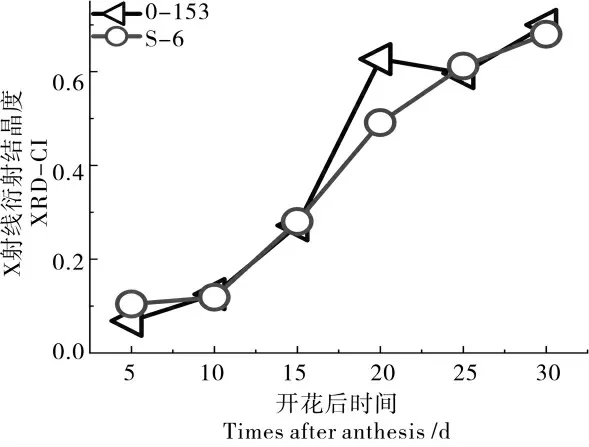
图5 0-153 和S-6 的XRD-CI 变化曲线Fig. 5 Changing of XRD-CI values of 0-153 and S-6 with fiber development
2.5 相关性分析
图6 所示为不同方法得到的FTIR-CI 与XRD-CI 的回归拟合相关性。 从决定系数R2可以看出, 最大为0.97,最小只有0.24,其中在0.9以上的有FTIR-CCI 法和陆地棉0-153 的FTIR-LOI 法。

图6 4 种计算方法得到的0-153 和S-6 棉纤维FTIR-CI 与XRD-CI 的相关性Fig. 6 Coefficient between FTIR-CI calculated by different methods and XRD-CI of 0-153 and S-6 fibers
2 个品系之间使用同一种FTIR-CI 与XRD-CI 拟合,所得到的决定系数R2接近,说明同一种方法在不同品系间拟合程度相当。
对于FTIR-TCI 方法,2 个品系的FTIR-CI 与XRD-CI 回归拟合相关性都较差。 对于FTIR-LOI方法,虽然陆地棉0-153 拟合R2超过0.9,海岛棉S-6 的R2也超过0.8,但该值逐渐减小。FTIR-HBI与XRD-CI 的决定系数R2分别为0.72 和0.77。对于FTIR-CCI,2 个材料决定系数R2都高于0.9。综上所述,FTIR-CCI 可以用于研究棉花纤维发育过程中结晶度的变化, 而FTIR-TCI、FTIR-HBI 方法可能不适合。
2.6 成熟纤维FTIR-CCI 分析
图7 所示为陆地棉0-153 和海岛棉S-6 合并后的FTIR-CCI 与XRD-CI 的回归拟合结果,R2为0.90,所得到的红外显微光谱(FTIR-CCI,x)的拟合模型结晶度 (IR-CI,y) 算法方程为y=3.71x-3.33,斜率标准误差为0.38,截距标准误差为0.39。

图7 0-153 和S-6 棉纤维FTIR-CCI 与XRD-CI 关联性Fig. 7 Total coefficient between FTIR-CCI and XRD-CI of 0-153 and S-6 fibers
由笔者使用相同的红外设备和XRD 衍射仪以及相同的校准方法在在相似的环境下进行实验操作, 分别获取18 个品系成熟纤维的红外显微光谱和XRD 图谱。 XRD 结果可以用作接受参照值。 红外显微光谱测量对于每个品系都选择至少10 个点, 测量后获取均值计算FTIR-CCI,将其代入模型获得IR-CI。 XRD-CI 和IR-CI 法的计算结果见表2。为明确本方法的精密度,计算得到测定结果的相对标准偏差,最大为0.123,最小为0.036,平均为0.079。 XRD-CI 和IR-CI 结果分布见图8。 如图8a,XRD-CI 结果的均值和中位数都比IR-CI 结果大,XRD-CI 值分布集中,IR-CI值的范围广。图8b 中XRD-CI 和IR-CI 所有值都分布于0.60~0.85。2 种方法的均方误差为0.038,XRD-CI 结果都在拟合模型计算结果的误差范围内。 使用该拟合模型可以评估成熟纤维结晶度,但是相比于发育中纤维,结果的偏差偏大。 因此,在下一步工作中使用更多的成熟纤维样品进行测试,进一步完善拟合模型。

表2 成熟纤维方法验证Table 2 Mature fiber method validation

图8 成熟纤维XRD-CI 与IR-CI 结果Fig. 8 The CI results of XRD and the model for mature fiber
3 讨论
本研究使用傅里叶变换红外显微光谱结合XRD 技术对2 个品系的发育中棉纤维的结晶度进行了一系列研究。 相比于传统的红外光谱仪,傅里叶变换红外显微光谱仪无色散原件,基于干涉调频分光,使物质的成分检测更方便快速。 红外光谱在理论上可以收集所有能够引发红外吸收的物质的振动信息,但部分红外信号太弱不会被仪器识别,同时复合物成分复杂,不同物质的红外峰可能发生部分重叠。 XRD 对于研究结晶材料有显著作用,可以获取结晶材料中结晶成分的空间分布等信息, 但是不能获取物质振动信息,也存在结晶成分与非晶成分的衍射峰重叠现象。 因此, 将傅里叶变换红外显微光谱与XRD联用, 可以快速获取物质的成分结构及结晶分布信息。
已有的4 种红外CI 计算方法, 都是使用纯度较高的纤维素建立的,并未直接用于自然棉花纤维。 本研究将文献中的4 种FTIR-CI 方法用于棉纤维的结晶度计算。 结果表明,FTIR-CCI 法适合研究棉纤维在发育时期的结晶度变化。 因此,利用FTIR-CCI 法结晶度结果与XRD-CI 结晶度结果拟合,建立模型。笔者尝试使用这个模型评估成熟纤维的结晶度。结果表明,该模型对于评估成熟棉纤维的结晶度效果有待进一步改进。 经分析,导致这一现象的可能原因如下:不同品系材料中, 由于成熟纤维各部位的发育程度不一致,而红外测试时的纤维为随机选取,导致其测试结果有偏差;在不同品系材料中,其成分分布、发育状态也不一致,这也会造成异常值的出现。
在下一步工作中,基于2 种方法中不同物质谱峰重叠现象,笔者将使用曲线拟合的方法尽可能将不同成分分离开来,获取更准确的结晶度信息。 对于成熟纤维,笔者将选择更多材料进行研究,同时缩小材料品系范围,增加每个材料测试的光谱数量,建立更适合评估成熟棉花纤维结晶度的FTIR 模型。
4 结论
使用micro-FTIR 对2 个棉花品系 (陆地棉0-153 和海岛棉S-6)在发育期纤维化学成分和结晶度进行了研究。 同时得到2 个材料随着开花后时间变化的XRD 谱图,并将FTIR-CI 与XRD-CI作回归拟合相关性分析。
通过对各个时期红外峰强度的比较,2 个材料总体趋势相同。 与FTIR-CI 相关的峰都逐渐增加,与非纤维素类物质如果胶等相关的峰则逐渐减小甚至消失。 通过红外峰和XRD 衍射峰强度比较以及XRD 结晶度变化趋势可以推测, 相比于海岛棉S-6,陆地棉纤维0-153 在开花后5~15 d 的发育慢,在开花后15~20 d 的发育快,开花后20 d 以后发育速率则保持相近。
4 种FTIR-CI 方法与XRD-CI 方法结果的回归拟合相关性结果表明, 使用FTIR-CCI 法可以很好地研究棉纤维发育阶段的结晶度变化。 同时, 采用该IR-CI 模型对成熟棉花纤维的结晶度进行计算,结果表明,该模型用于评估成熟棉花纤维的结晶度的精密度还有待提高,需要使用更多的成熟纤维建立优化的模型。
猜你喜欢
纺织科技进展(2021年4期)2021-07-22
纺织科技进展(2021年3期)2021-06-09
纺织科技进展(2021年3期)2021-06-09
陶瓷学报(2021年1期)2021-04-13
今日农业(2020年17期)2020-12-15
今日农业(2020年14期)2020-12-14
现代农村科技(2020年9期)2020-09-08
丝绸(2020年6期)2020-06-23
云南农业科技(2019年2期)2019-04-04
江苏农业科学(2015年1期)2015-04-17
