
盐胁迫下粳稻品种南粳9108分蘖特性及其与群体生产力的关系
2020-07-21 08:29韦还和葛佳琳张徐彬孟天瑶李心月丁恩浩陈英龙戴其根
作物学报 2020年8期
韦还和 葛佳琳 张徐彬 孟天瑶 陆 钰 李心月 陶 源 丁恩浩 陈英龙 戴其根,*
盐胁迫下粳稻品种南粳9108分蘖特性及其与群体生产力的关系
韦还和1葛佳琳1张徐彬1孟天瑶2陆 钰1李心月1陶 源1丁恩浩1陈英龙1戴其根1,*
1江苏省作物遗传生理重点实验室 / 江苏省作物栽培生理重点实验室 / 江苏省粮食作物现代产业技术协同创新中心 / 扬州大学水稻产业工程技术研究院, 扬州大学, 江苏扬州 225009;2扬州大学教育部农业与农产品安全国际合作联合实验室 / 扬州大学农业科技发展研究院, 扬州大学, 江苏扬州 225009
旨在明确盐胁迫下水稻分蘖发生与成穗规律及其优势叶位, 为沿海滩涂水稻高产栽培的分蘖合理利用与调控提供依据。以江苏沿海滩涂大面积种植的常规粳稻南粳9108为试材, 设置对照(盐浓度0)、中盐(盐浓度0.15%)和高盐(盐浓度0.3%)环境, 比较研究盐逆境下水稻分蘖发生与成穗特性及其对群体产量的贡献。结果表明, 对照、中盐和高盐处理的两年平均产量(t hm–2)分别为9.7、7.4和4.2; 中盐和高盐处理的穗数、每穗粒数、结实率和千粒重均显著低于对照。与对照相比, 中盐和高盐处理下拔节、抽穗和成熟期群体茎蘖数和成穗率均较低。对照的分蘖利用以一次分蘖和二次分蘖为主, 一次分蘖发生在第3至第7叶位, 第4至第6叶位是分蘖发生与成穗的优势叶位, 二次分蘖则以1/4和1/5蘖位优势较强; 盐胁迫的分蘖利用以一次分蘖为主, 第4至第6叶位是分蘖发生与成穗的优势叶位。盐胁迫下各蘖位的穗长、每穗粒数、着粒密度、一次枝粳数及粒数、二次枝粳数及粒数均低于对照。与对照相比, 盐胁迫下水稻单株成穗数少、个体和群体生长协调性差、穗型小, 最终单株和群体产量低。
盐胁迫; 水稻; 分蘖特性; 群体生产力
水稻是我国主要的口粮作物, 在粮食安全中起着举足轻重的作用[1]。当前, 我国粮食需求迅速增长、耕地面积不断减少、淡水资源日益稀缺等多重挑战, “人、地、粮”矛盾依旧突出。我国有234万公顷沿海滩涂和1亿公顷内陆盐碱地, 是我国最重要的后备耕地资源, 综合利用潜力巨大[2-3]。盐胁迫对水稻生长的影响一直备受关注, 国内外学者已在盐胁迫下水稻种子萌发、幼苗生长发育、光合作用与干物质积累以及产量构成等方面开展了一系列研究[4-6]。盐胁迫使叶绿体结构破坏, 叶绿素含量降低, 叶片净光合速率下降, 抑制水稻植株光合作用和同化物积累[4]。水稻常作为盐土改良的重要粮食作物, 我国各地的沿海滩涂和内陆盐碱地均在推广种植水稻[7-8]。但总体而言, 沿海滩涂和内陆盐碱地种植的水稻产量偏低且稳产性较差[9-10], 一个重要原因是对盐胁迫下水稻产量形成生理特性的研究尚不够深入。单位面积穗数是水稻产量构成要素之一, 主要取决于分蘖发生数量和成穗率[11]。分蘖是影响水稻穗数进而决定水稻产量的重要农艺性状, 水稻分蘖性强弱不仅与自身遗传特性相关, 而且也受农艺措施和栽培环境重要影响[12-13]。已有研究表明, 栽插密度、水肥管理、机械作业方式等均可影响水稻分蘖发生与成穗[14-16]; 光照和温度等也影响水稻分蘖发生与成穗[17-18]。目前, 关于沿海滩涂水稻的分蘖发生与成穗特点的研究较少, 且缺少系统报道。本文研究盐胁迫下水稻分蘖发生与成穗特点, 以期明确沿海滩涂水稻分蘖发生与成穗规律及优势叶位, 为江苏沿海滩涂水稻高产栽培的分蘖合理利用与调控措施提供依据。
1 材料与方法
1.1 供试材料
以常规粳稻南粳9108为试材, 主茎总叶数为16叶, 伸长节间数为6个。
1.2 试验设计与栽培管理
试验于2018—2019年在扬州大学农学院试验农场盐池进行, 共6个盐池, 每个盐池内长3.6 m、内宽1.5 m、内深0.5 m。盐池内土壤类型为沙壤土, 含全氮0.14%、碱解氮92.4 mg kg–1、速效磷33.8 mg kg–1、速效钾83.1 mg kg–1。试验设置3个盐浓度梯度处理, 即对照(Control, CK, 盐浓度0)、中盐(Medium-salinity stress, MS, 盐浓度0.15%)和高盐(High-salinity stress, HS, 盐浓度0.3%), 每处理小区面积为5.4 m2(3.6 m×1.5 m), 2次重复。按土壤体积、密度和盐浓度计算出各小区应施用的海盐重量(选用浙江蓝海星盐制品厂生产的Q/ZLY型速溶海水晶), 并在水稻秧苗移栽前3 d, 将海盐撒施至对应盐池中, 充分搅拌均匀。盐池上方建有遮雨棚, 以防降雨引起池内盐分变化。
采用塑料软盘旱育秧, 于5月23日播种, 6月13日移栽。移栽叶龄3.8叶左右, 无带株分蘖。栽插密度为28.6万穴 hm–2(25 cm×14 cm), 每穴3株苗栽插。总施氮量270 kg hm–2, 基蘖肥∶穗肥 = 6∶4, 其中基肥在移栽前1 d施用, 分蘖肥于移栽后10 d施用, 穗肥于倒四叶、倒二叶等量施用。过磷酸钙(含12% P2O5) 1125 kg hm–2, 作基肥一次施用; 钾肥50%作为基肥, 50%于倒四叶施用。秧苗移栽后采用湿润灌溉; 有效分蘖临界叶龄期的前1个叶龄开始控水搁田; 拔节至成熟期实行干湿交替灌溉。按常规高产栽培要求防治病虫草害。
1.3 测定项目与方法
1.3.1 分蘖发生与成穗的追踪 从每个小区中心区域选取长势比较一致连续3行、每行6穴的植株。每5 d一次标记叶龄, 并对分蘖的发生情况挂牌追踪标志。为每个主茎和分蘖挂上标签, 在标签上写好分蘖的次级和叶位。成熟期根据各分蘖的标记牌将各级分蘖分开, 单独收获, 记录各级各叶位分蘖的发生数、成穗数, 然后单独考种, 分穗长、一次枝粳数、二次枝粳数等考察穗部性状。
各蘖位分蘖的发生率指该蘖位分蘖的实际发生数量与观察株数的百分比, 各蘖位分蘖的成穗率指该蘖位分蘖的成穗数与分蘖实际发生数的百分比。X/0分蘖指着生在主茎第X叶位上的一次分蘖, X/1分蘖指主茎第1叶位一次分蘖的第X叶位上的二次分蘖, 如1/1分蘖为主茎第1叶位一次分蘖的第1叶位上的二次分蘖, 依此类推。
1.3.2 成熟期干物质和产量测定 于成熟期, 按各小区茎蘖数的平均数取5穴, 测定干物质量。各小区调查60穴, 计算有效穗数; 取5穴调查每穗粒数、结实率和测定千粒重及理论产量, 小区实收测产。
1.4 数据处理
运用Microsoft Excel软件录入数据、计算, 用DPS软件作统计分析。由于2年试验结果趋势基本一致, 本文以2年数据平均值进行分析, 仅在1年试验中出现的分蘖, 按出现年度计算平均值。
2 结果与分析
2.1 产量与干物质
两年中, 对照、中盐和高盐处理的平均产量(t hm–2)分别为9.7、7.4和4.2。与对照相比, 中盐和高盐处理的产量降幅分别为23.7%和56.7%。产量构成因素方面, 中盐和高盐处理的穗数和每穗粒数均低于对照, 差异显著; 颖花量较对照分别低20.7%和48.0% (两年平均值)。就结实率和千粒重而言, 中盐和高盐处理下均低于对照; 两年中, 中盐和高盐处理下的千粒重(g)分别为22.9和21.4, 较对照分别低1.1 g和2.6 g (表1)。中盐和高盐处理的成熟期干物重显著低于对照, 两年平均较对照分别低26.9%和60.0%; 收获指数则以中盐和高盐处理高于对照(表1)。
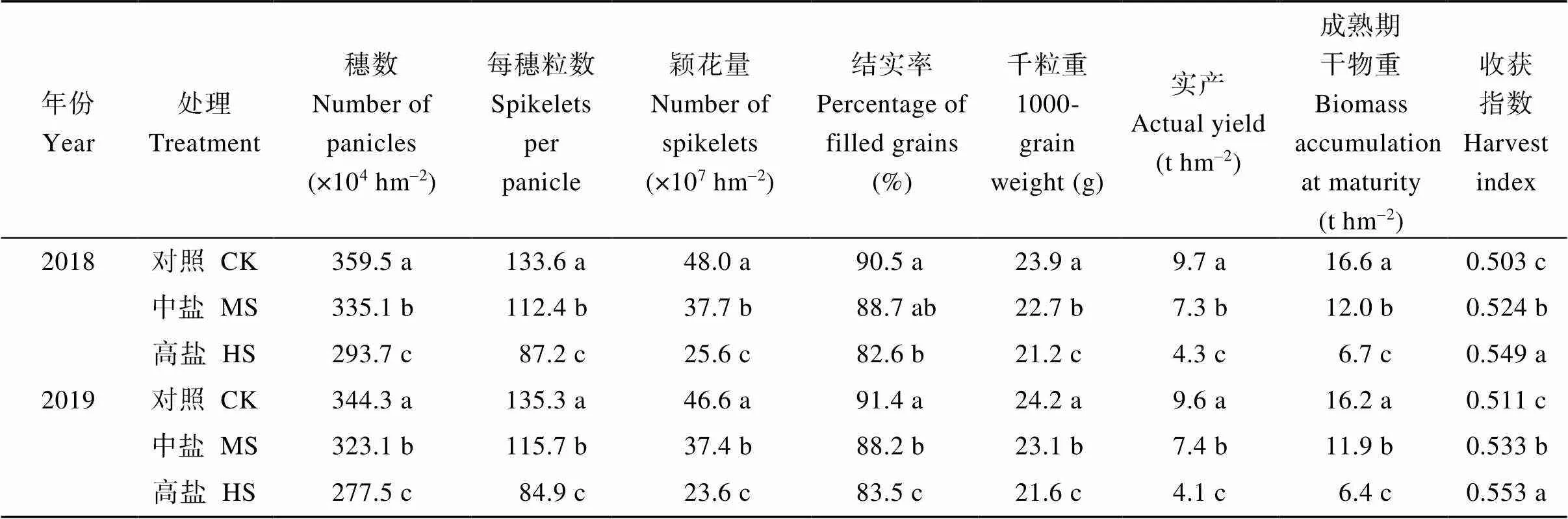
表1 盐胁迫对水稻产量及其构成因素的影响
CK: 对照; MS: 中盐; HS: 高盐。标以不同小写字母的值在同一年份同一列中5%水平差异显著。
CK: control; MS: medium-salinity stress; HS: high-salinity stress. Values within the same column and the same year followed by different lowercase are significantly different at the 5% probability level.
2.2 关键生育期群体茎蘖数
不同处理之间在拔节、抽穗和成熟期的群体茎蘖数差异均显著(表2)。两年中, 对照、中盐和高盐处理拔节期的群体茎蘖数(×104hm–2)分别为454.6、441.4和396.6, 高盐处理下拔节期的茎蘖数较对照低12.8%。抽穗期对照、中盐和高盐处理的群体茎蘖数(×104hm–2)分别为358.2、345.3和305.4, 中盐和高盐处理下的茎蘖数较对照分别低3.6%和14.7%。成熟期对照、中盐和高盐处理的群体茎蘖数(×104hm–2)分别为351.9、329.1和285.6, 中盐和高盐处理下的茎蘖数较对照分别低6.5%和18.8% (表2)。各处理自拔节期后的群体茎蘖数均下降, 但以对照处理降幅最小、高盐处理降幅最大, 最终成穗率以对照最高、高盐处理最低; 两年中, 对照的成穗率(%)为77.4, 显著高于中盐(74.6%)和高盐(72.0%)处理(图1)。
2.3 分蘖发生叶位及发生率
分蘖发生率方面, 对照一次分蘖叶位有3/0至7/0共5个, 二次分蘖叶位有1/4、2/4、3/4、1/5、2/5和1/6共6个。中盐处理一次分蘖叶位和二次分蘖叶位均与对照一致。高盐处理一次分蘖叶位有4/0至7/0共4个, 二次分蘖叶位有1/4、2/4、1/5和2/5共4个。一次分蘖4/0至7/0的分蘖发生率均呈对照>中盐>高盐, 二次分蘖1/4、2/4、1/5和2/5亦呈现类似趋势(表3)。分蘖成穗率方面, 对照一次分蘖以4/0、5/0、6/0分蘖成穗率较高, 均在70%以上; 二次分蘖除2/5叶位分蘖未成穗外, 其余叶位分蘖均有成穗。中盐和高盐处理一次分蘖同样以4/0、5/0、6/0分蘖成穗率较高; 中盐处理二次分蘖以3/4和2/5叶位未能成穗, 1/4、2/4、1/5和1/6叶位均有成穗; 高盐处理二次分蘖则以2/4、3/4、2/5和1/6叶位未能成穗, 仅1/4和1/5叶位有成穗(表3)。
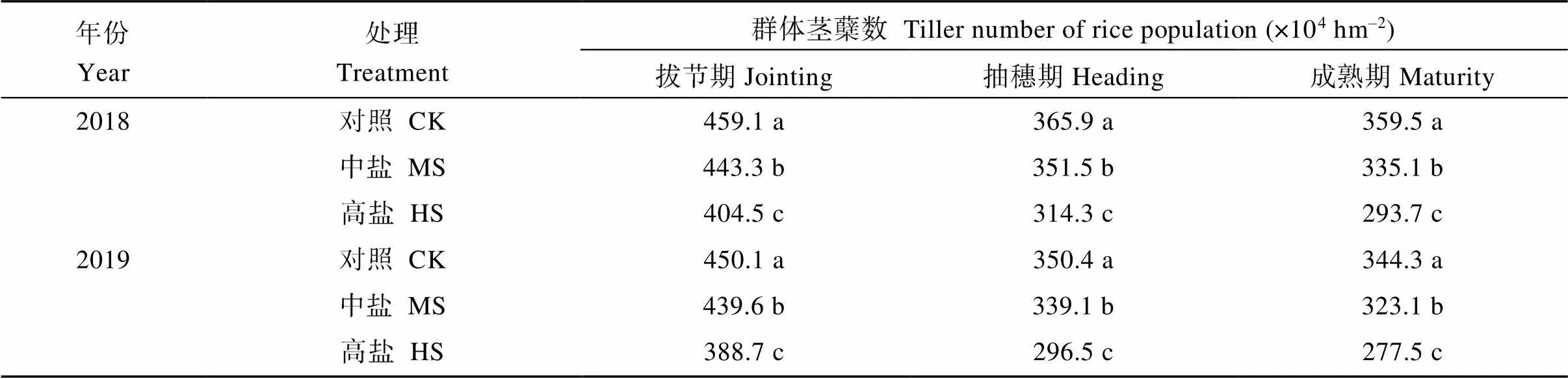
表2 盐胁迫对水稻关键生育期群体茎蘖数的影响
CK: 对照; MS: 中盐; HS: 高盐。标以不同小写字母的值在同一年份同一列中5%水平差异显著。
CK: control; MS: medium-salinity stress; HS: high-salinity stress. Values within the same column and the same year followed by different lowercase are significantly different at the 5% probability level.
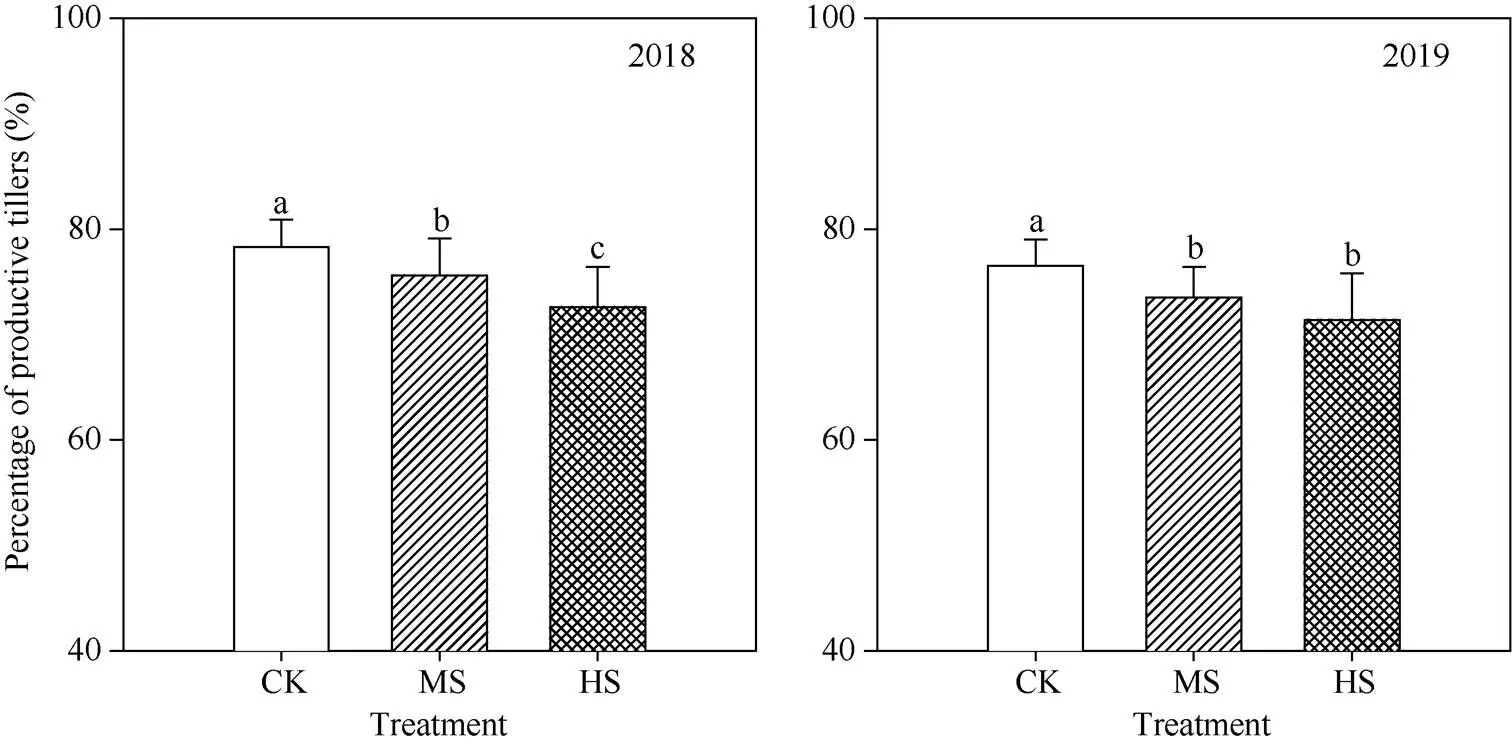
图1 盐胁迫对水稻成穗率的影响
CK: 对照; MS: 中盐; HS: 高盐。处理间标以不同小写字母的值在5%水平差异显著。
CK: control; MS: medium-salinity stress; HS: high-salinity stress. Bars superscripted by different lowercase are significantly different at the 5% probability level between treatments.
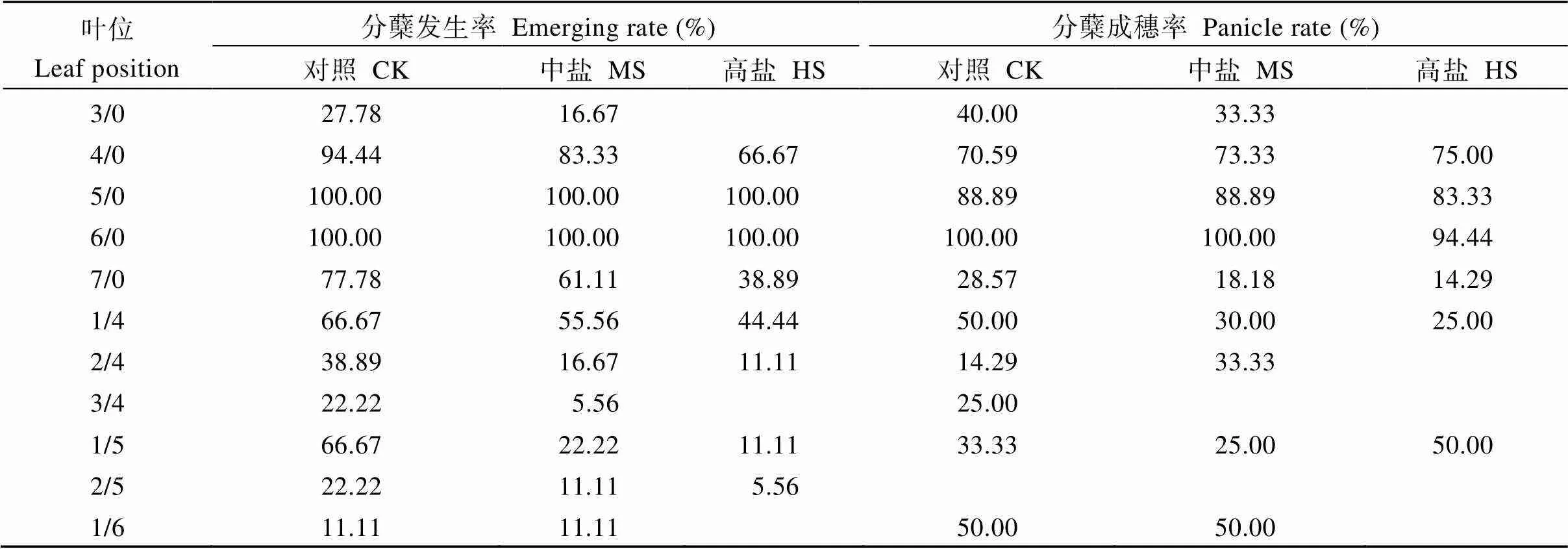
表3 盐胁迫对水稻分蘖发生及成穗率的影响
CK: 对照; MS: 中盐; HS: 高盐。
CK: control; MS: medium-salinity stress; HS: high-salinity stress.
2.4 单株成穗数的茎蘖组成及对产量的贡献
由表4可知, 对照的单株成穗数为4.61, 显著高于中盐和高盐处理。对照的一次分蘖数和二次分蘖数亦均高于中盐和高盐处理。对照单株成穗数一次分蘖以4/0至6/0为主, 二次分蘖则以1/4和1/5为主; 中盐和高盐处理单株成穗数则主要依靠主茎和4/0至6/0叶位的一次分蘖(表4)。
与茎蘖组成相类似, 对照群体产量显著高于中盐和高盐处理; 此外, 主茎、一次分蘖和二次分蘖叶位的产量亦以对照>中盐>高盐。就产量贡献率而言, 对照一次分蘖以4/0至6/0叶位贡献较大, 二次分蘖则以1/4和1/5叶位贡献较大; 中盐和高盐处理下群体产量贡献率主要依靠主茎和4/0至6/0叶位的一次分蘖(表5)。
2.5 各叶位茎蘖的穗部性状
各处理在穗部主要性状指标上差异较明显。各处理叶位的穗长、总粒数、结实率、着粒密度、一次枝粳数及粒数、二次枝粳数及粒数的平均值均呈对照>中盐>高盐。各处理共有的一次分蘖4/0至6/0的穗长、总粒数、着粒密度、一次枝粳数及粒数、二次枝粳数及粒数也以对照最高、高盐处理最低; 共有的二次分蘖1/4和1/5的穗部性状指标值亦呈上述趋势(表6)。
3 讨论
3.1 盐胁迫下水稻分蘖发生与成穗特点
水稻的分蘖与成穗特点既受自身遗传特性调控,又易受外界环境的影响[19]。在适宜的条件下, 水稻分蘖按N–3的规律与主茎叶同伸, 促使分蘖早生快发, 充分利用有效蘖位, 进而提高茎蘖成穗率是水稻高产群体的共性特征[20]。本研究结果表明, 与对照相比, 盐逆境下水稻低位分蘖缺位较多。一方面是由于机插软盘育秧的播量密度大引起叶蘖不同伸[11,21]; 另一方面, 由于移栽(三叶一心期移栽)植伤较重, 小苗抗盐逆境较弱, 导致低位分蘖缺位[22]。分蘖发生率方面, 对照一次分蘖叶位共有5个, 二次分蘖叶位共有6个; 而高盐处理一次分蘖和二次分蘖叶位均为4个。分蘖成穗率方面, 盐胁迫下水稻一次分蘖与对照差异较小, 而二次分蘖则与对照差异较大。与对照相比, 盐逆境下水稻一次分蘖和二次分蘖的发生数低, 且分蘖成穗率尤其是二次分蘖成穗率也较低, 造成单株成穗数以及群体穗数均较低。
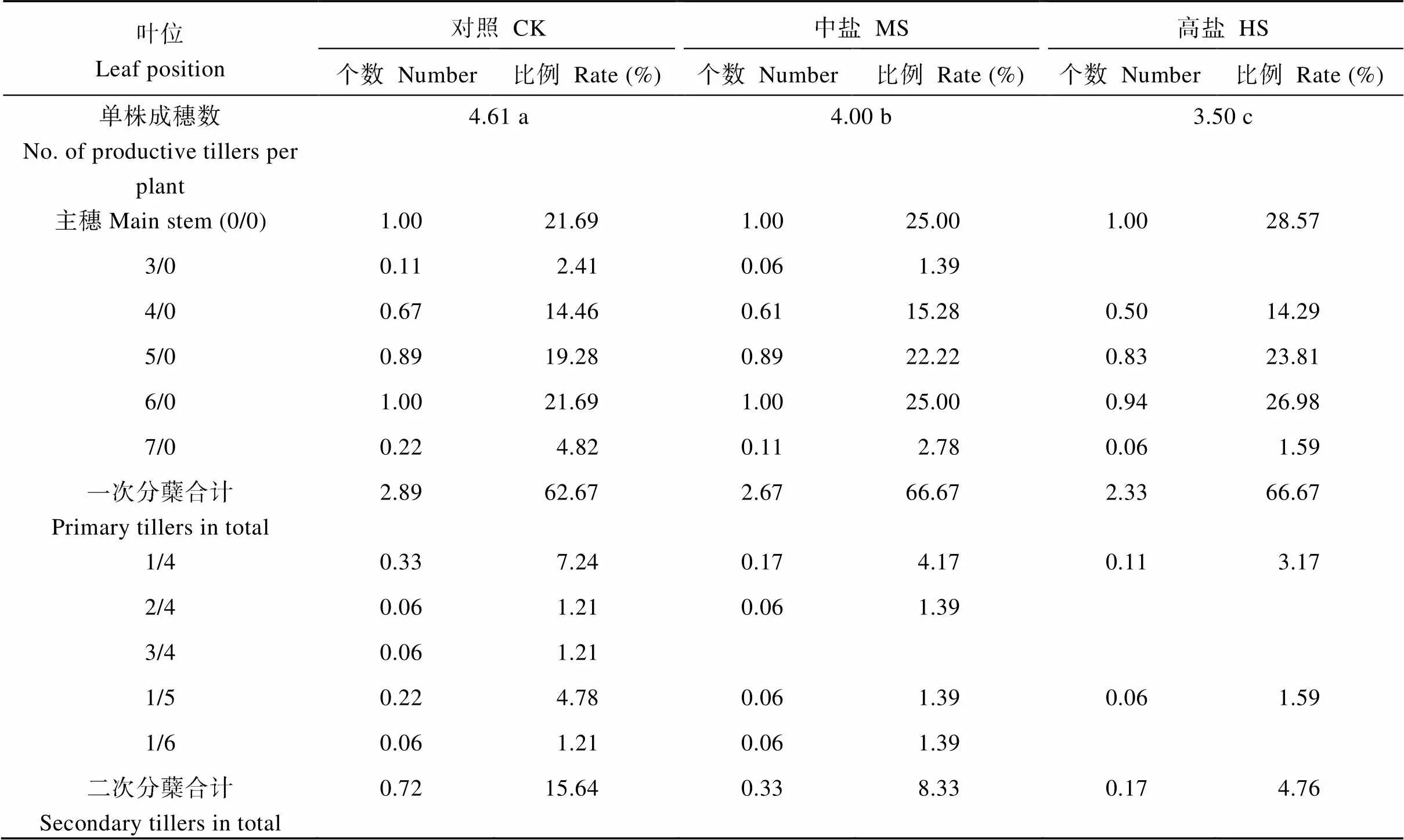
表4 盐胁迫下水稻单株成穗数的茎蘖组成
CK: 对照; MS: 中盐; HS: 高盐。标以不同小写字母的值在5%水平差异显著。
CK: control; MS: medium-salinity stress; HS: high-salinity stress. Values followed by different lowercase are significantly different at the 5% probability level.
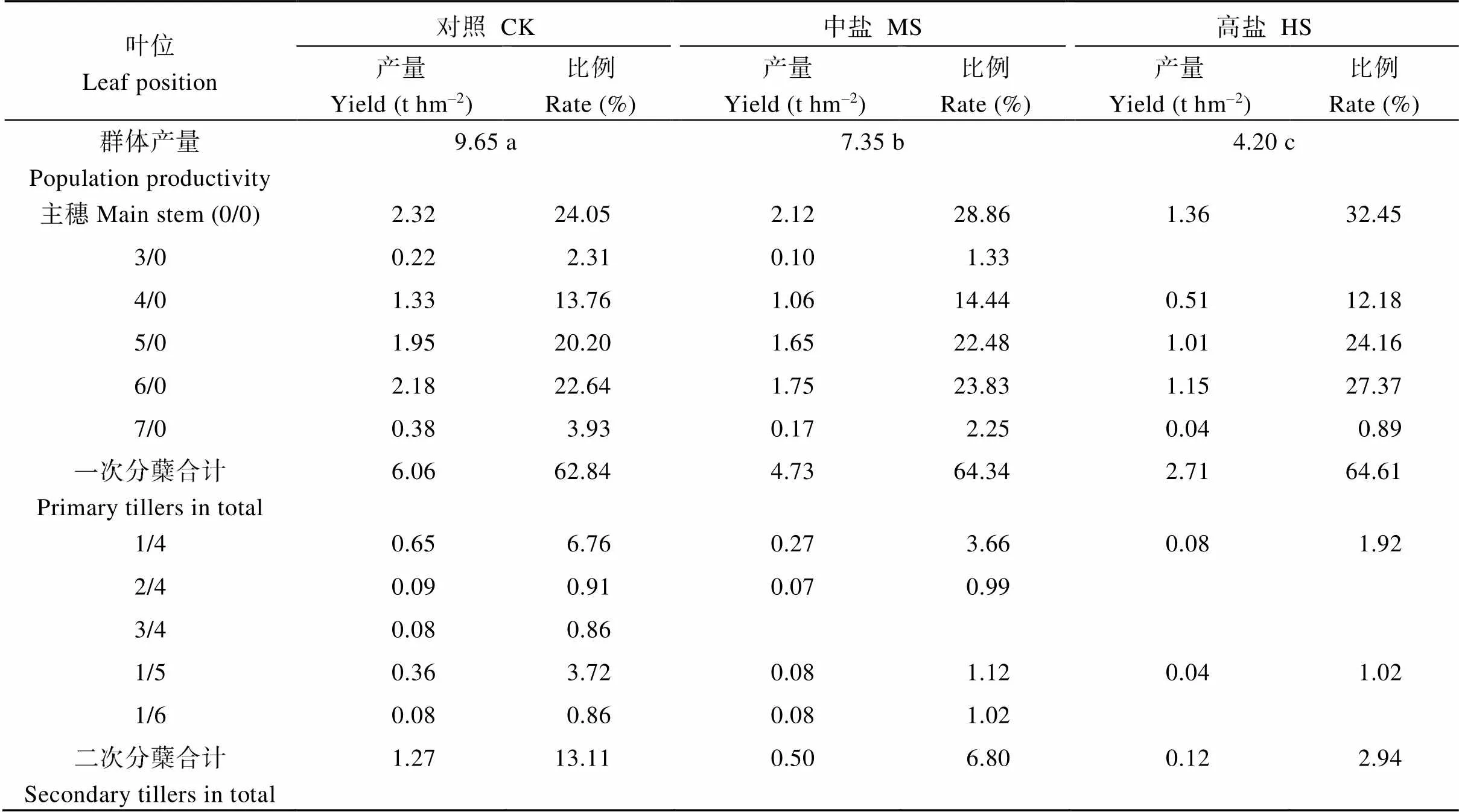
表5 盐胁迫下水稻各叶位茎蘖对群体产量的贡献
CK: 对照; MS: 中盐; HS: 高盐。标以不同小写字母的值在5%水平差异显著。
CK: control; MS: medium-salinity stress; HS: high-salinity stress. Values followed by different lowercase are significantly different at the 5% probability level.
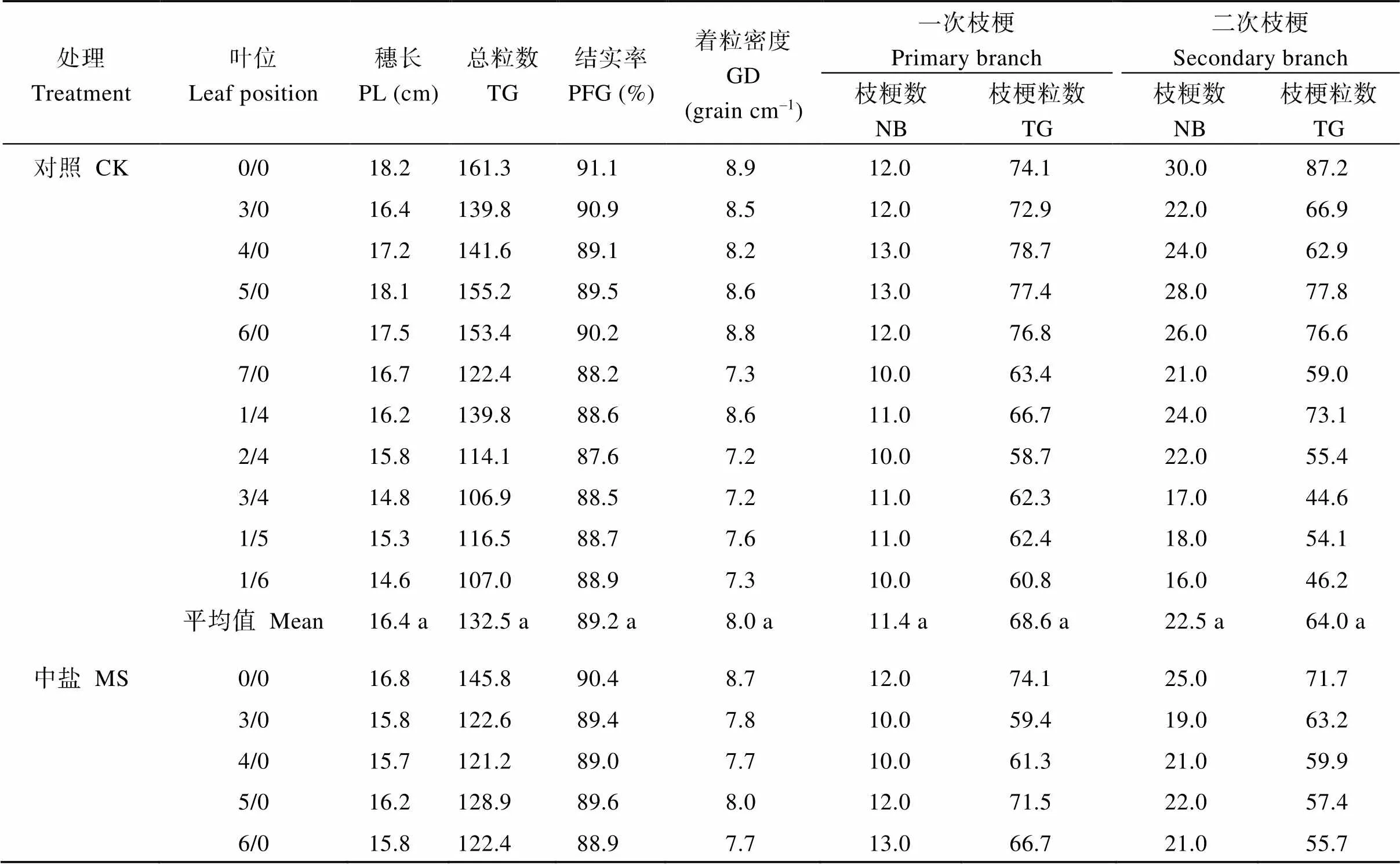
表6 盐胁迫对水稻各叶位茎蘖穗部性状的影响
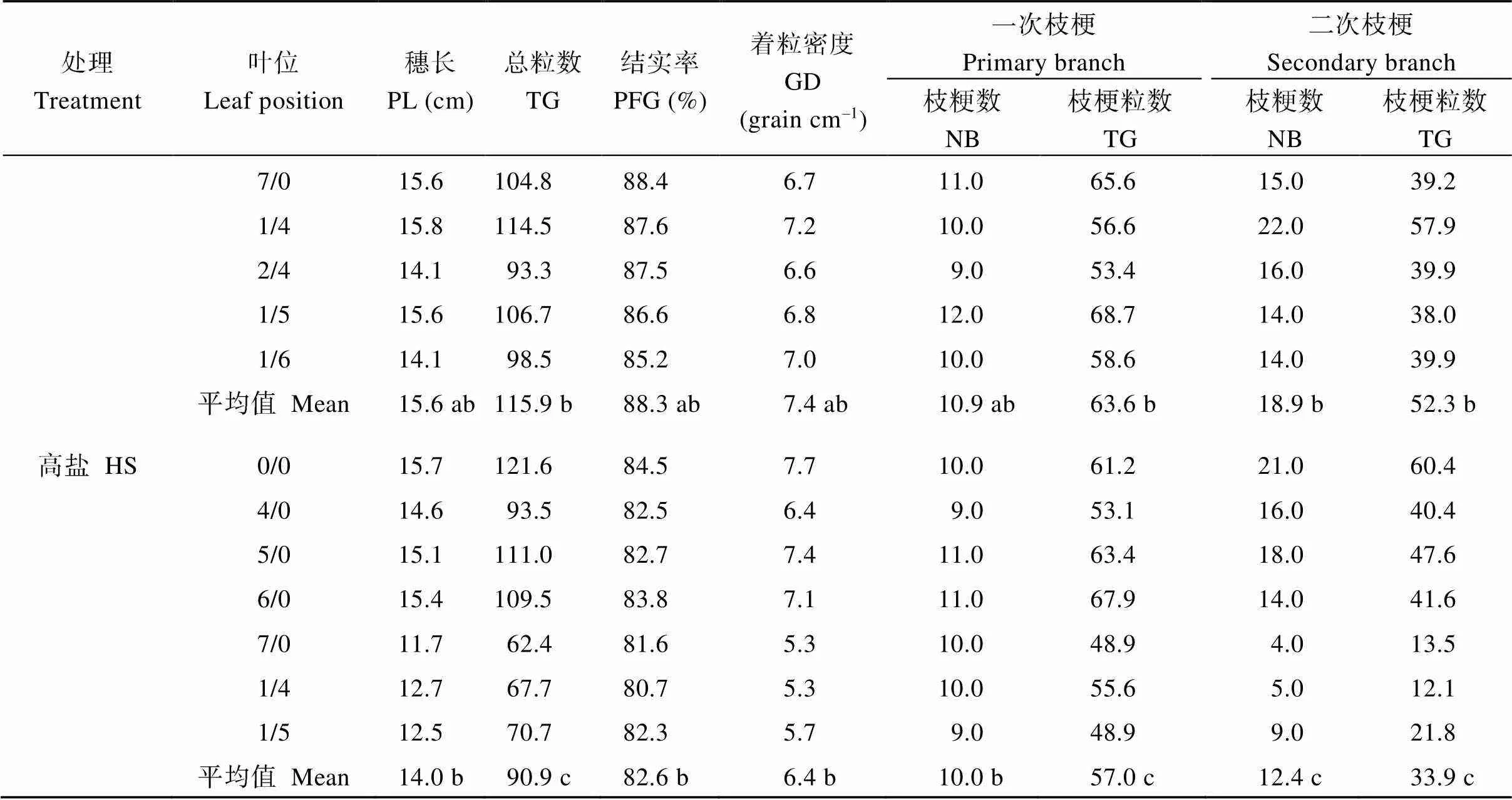
(续表6)
CK: 对照; MS: 中盐; HS: 高盐。标以不同小写字母的值在5%水平差异显著。
CK: control; MS: medium-salinity stress; HS: high-salinity stress. PL: panicle length; TG: total grains; PFG: percentage of filled grains; GD: grain density; NB: number of branches. Values followed by different lowercase are significantly different at the 5% probability level.
成穗率是表征群体质量的重要指标。一般而言, 在群体干物质积累量相近的情况下, 群体茎蘖成穗率越高, 作物群体产量也越高[23-24]。本试验条件下, 与对照相比, 盐胁迫下拔节、抽穗和成熟期的群体茎蘖数均较低, 且拔节至成熟阶段的群体茎蘖数消退幅度大, 最终成穗率和群体产量也较低。因此, 在沿海滩涂水稻生产中, 在保证适宜穗数的前提下, 应注重提高群体的茎蘖成穗率, 从而优化群体质量。
3.2 盐胁迫下水稻优势蘖位利用及其对产量的贡献
水稻分蘖发生与成穗普遍存在优势叶位现象, 充分利用和提高优势蘖位的比值, 利于优化群体质量和提高产量[25-26]。目前, 有关不同基因型和栽培方式下水稻优势叶位的研究已有较多报道。基因型方面, 如吕伟生等[11]研究表明, 早稻分蘖发生和成穗的优势叶位在主茎第3至第6叶位的一次分蘖, 其中常规稻的优势叶位为主茎第4至第5叶位, 杂交稻则在主茎第3至第5叶位; 栽培方式方面, 如李杰等[26]研究表明, 手栽稻优势叶位在第5至第8叶位的一次分蘖和1/5、1/6和1/7叶位的二次分蘖; 机插稻优势叶位在第4至第7叶位的一次分蘖和1/4叶位的二次分蘖; 直播稻优势叶位则在第1至第4叶位的一次分蘖。本试验条件下, 对照优势叶位为4/0至6/0叶位的一次分蘖和1/4、1/5叶位的二次分蘖, 而中盐和高盐处理优势叶位则为4/0至6/0叶位的一次分蘖; 此外, 盐胁迫下4/0至6/0叶位一次分蘖的穗长、每穗粒数、着粒密度、一次枝粳数及粒数、二次枝粳数及粒数亦均高于其余蘖位。本研究结果表明, 中部4/0至6/0叶位一次分蘖的分蘖发生与成穗率高、穗部性状好, 产量贡献率也较大, 是盐胁迫下水稻产量形成的基础。
前人研究表明[3,5,8], 不同水稻品种对盐胁迫的耐受性存在较大差异。本研究受限于盐池的数量和面积, 在品种选择上以当前江苏沿海滩涂大面积种植的常规粳稻南粳9108为试材, 明确了盐胁迫下其分蘖发生与成穗规律, 研究结果对于指导盐碱地水稻品种的选育和栽培调控具有一定意义。本研究部分研究结果仍有必要进一步在实际滩涂盐地以及耐盐性不同的品种中验证。
3.3 沿海滩涂水稻高产群体分蘖合理利用与调控的关键栽培技术
已有的研究报道和生产实际表明, 库容量偏小以及库容充实度差限制了沿海滩涂水稻产量的提升[27-29]。本试验条件下, 中盐和高盐处理的库容量(群体颖花量)和库容充实度(结实率和千粒重)均低于对照, 最终产量较对照分别降低23.7%和56.7%, 这与前人的研究结果一致[3,8,27-29]。盐胁迫下机插软盘小苗盐逆境抗性较弱, 移栽后植伤较重, 导致低位分蘖缺位[21-22]; 此外, 盐胁迫下植株光合作用受到抑制, 光合产物积累量少[3-4], 加剧了光合产物在主茎和分蘖之间的竞争, 引起部分分蘖因难以获得充足的养分最终未能成穗, 较低的分蘖发生率以及分蘖成穗率导致盐胁迫下水稻群体穗数的降低。在试验进程中, 作者观察到盐胁迫处理下的穗部颖花退化数明显高于对照(数据尚未发表), 这也是盐胁迫下每穗粒数较低的一个重要原因。因此, 盐胁迫下水稻穗数和每穗粒数均较低, 导致水稻库容量偏低。而在库容充实方面, 由于盐胁迫下植株光合作用和同化物积累受到抑制, 花后植株的衰老进程大为加快, 花后干物质积累量少[3-5,8], 导致结实率和千粒重偏低。
水稻分蘖主要发生在生育前期, 该时期合理群体的构建不仅对形成适宜穗数和扩大库容特别重要, 而且对于灌浆阶段库容的充实也至关重要[30]。针对江苏沿海滩涂水稻种植以常规粳稻为主的实际[3,31], 围绕常规粳稻品种的分蘖特性以及适应机械化、高效化生产的要求, 栽培上应注意如下几点。(1)确定适宜播种量, 稀播匀播。播种量过大是制约生产上水稻壮秧培育与大田高产群体起点建立的突出原因[32], 因此, 合理降低播量是关键, 并强调机械匀播, 依此提高秧苗素质和培育带蘖壮秧, 促进穴间均衡生长。(2)适当增加基本苗。针对沿海滩涂水稻栽后易死苗、僵苗[33]以及4/0至6/0一次分蘖的优势蘖位, 应适当增加栽插基本苗, 利于在有效分蘖期充分发挥优势分蘖生长潜力, 形成足量的壮个体, 又可在无效分蘖期利用优势蘖位对无效分蘖进行生理调节, 形成合理的群体茎蘖动态, 提高成穗率和群体质量。(3)水分的合理调控。实践表明, 沿海盐土地种稻成功最基本的条件是引淡水灌溉洗盐[7], 灌水排盐也是沿海滩涂种植水稻的重要措施。在水分管理上, 尤其是搁田期要遵循“轻、多”的原则, 分多次自然断水轻搁田, 一方面促进有效分蘖正常生长和控制无效分蘖, 提高群体质量, 另一方面避免因土壤过度落干造成返盐, 影响植株正常生长。
4 结论
盐逆境对水稻分蘖发生与成穗规律有显著影响。对照分蘖利用以4/0、5/0、6/0一次分蘖和1/4、1/5二次分蘖为主, 盐胁迫下分蘖利用则以4/0、5/0、6/0一次分蘖为主。与对照相比, 盐胁迫下水稻单株成穗数少、个体和群体生长协调差、穗型小, 最终单株和群体产量低。沿海滩涂水稻生产中应根据盐胁迫下水稻分蘖发生与成穗特点以及品种特性, 在适播匀播稀播提高秧苗素质基础上, 适当增加大田基本苗, 并搭配水、肥合理农艺, 促进优势叶位分蘖早发、快发, 充分发掘优势叶位的增产潜力, 实现沿海滩涂水稻的高产高效生产。
[1] Liu L L, Zhu Y, Tang L, Cao W X, Wang E L. Impacts of climate changes, soil nutrients, variety types and management practices on rice yield in East China: A case study in the Taihu region., 2013, 149: 40–48.
[2] 王才林, 张亚东, 赵凌, 路凯, 朱镇, 陈涛, 赵庆勇, 姚姝, 周丽慧, 赵春芳, 梁文化, 孙明法, 严国红. 耐盐碱水稻研究现状、问题与建议. 中国稻米, 2019, 25(1): 1–6. Wang C L, Zhang Y D, Zhao L, Lu K, Zhu Z, Chen T, Zhao Q Y, Yao S, Zhou L H, Zhao C H, Liang W H, Sun M F, Yan G H. Research status, problems and suggestions on salt-alkali tolerant rice., 2019, 25(1): 1–6 (in Chinese with English abstract).
[3] 周根友, 翟彩娇, 邓先亮, 张蛟, 张振良, 戴其根, 崔士友. 盐逆境对水稻产量、光合特性及品质的影响. 中国水稻科学, 2018, 32: 146–154.Zhou G Y, Zhai C J, Deng X L, Zhao J, Zhang Z L, Dai Q G, Cui S Y. Performance of yield, photosynthesis and grain quality ofrice cultivars under salinity stress in micro- plots., 2018, 32: 146–154 (in Chinese with English abstract).
[4] Ashraf M, Athar H R, Harris P J C, Kwon T R. Some prospective strategies for improving crop salt tolerance., 2008, 97: 45–110.
[5] 孙现军, 姜奇彦, 胡正, 张惠媛, 徐长兵, 邸一恒, 韩龙植, 张辉. 水稻资源全生育期耐盐性鉴定筛选. 作物学报, 2019, 45: 1656–1663.Sun X J, Jiang Q Y, Hu Z, Zhang H Y, Xu C B, Di Y H, Han L Z, Zhang H. Screening and identification of salt-tolerant rice germplasm in whole period., 2019, 45: 1656–1663 (in Chinese with English abstract).
[6] 沙汉景, 胡文成, 贾琰, 王新鹏, 田雪飞, 于美芳, 赵宏伟. 外源水杨酸、脯氨酸和γ-氨基丁酸对盐胁迫下水稻产量的影响. 作物学报, 2017, 43: 1677–1688. Sha H J, Hu W C, Jia Y, Wang X P, Tian X F, Yu M F, Zhao H W. Effect of exogenous salicylic acid, proline, and γ-aminobutyric acid on yield of rice under salt stress., 2017, 43: 1677–1688 (in Chinese with English abstract).
[7] 凌启鸿. 盐碱地种稻有关问题的讨论. 中国稻米, 2018, 24(4): 1–2. Ling Q H. Discussion on the related problems of rice planting in saline-alkali soil., 2018, 24(4): 1–2 (in Chinese with English abstract).
[8] 徐晨, 凌风楼, 徐克章, 武志海, 刘晓龙, 安久海, 赵兰坡. 盐胁迫对不同水稻品种光合特性和生理生化特性的影响. 中国水稻科学, 2013, 27: 280–286.Xu C, Ling F L, Xu K Z, Wu Z H, Liu X L, An J H, Zhao L P. Effects of salt stress on photosynthetic characteristics and physiological and biochemical traits of different rice varieties., 2013, 27: 280–286 (in Chinese with English abstract).
[9] Zeng L H, Shannon M C. Effects of salinity on grain yield and yield components of rice at different seeding densities., 2000, 92: 418–423.
[10] Gregorio G B, Senadhira D, Mendoza R D, Manigbas N L, Roxas J P, Guerta C Q. Progress in breeding for salinity tolerance and associated abiotic stresses in rice., 2002, 76: 91–101.
[11] 吕伟生, 曾勇军, 石庆华, 潘晓华, 黄山, 商庆银, 谭雪明, 李木英, 胡水秀. 机插早稻分蘖成穗特性及基本苗公式参数研究. 作物学报, 2016, 42: 427–436.Lyu W S, Zeng Y J, Shi Q H, Pan X H, Huang S, Shang Q Y, Tan X M, Li M Y, Hu S X. Tillering and panicle formation characteristics of machine-transplanted early rice and its parameters of basic population formulae., 2016, 42: 427–436 (in Chinese with English abstract).
[12] 韦还和, 李超, 张洪程, 孙玉海, 孟天瑶, 杨筠文, 马荣荣, 王晓燕, 戴其根, 霍中洋, 许轲, 魏海燕. 水稻甬优12超高产群体分蘖特性及其与群体生产力的关系. 作物学报, 2014, 40: 1819–1829.Wei H H, Li C, Zhang H C, Sun Y H, Meng T Y, Yang J W, Ma R R, Wang X Y, Dai Q G, Huo Z Y, Xu K, Wei H Y. Tillering characteristics and its relationship with population productivity of super-high yield rice population of Yongyou 12., 2014, 40: 1819–1829 (in Chinese with English abstract).
[13] 宋云生, 张洪程, 戴其根, 杨大柳, 郭保卫, 朱聪聪, 霍中洋, 许轲, 魏海燕, 胡加敏, 吴爱国, 蒋晓鸿. 水稻机栽钵苗单穴苗数对分蘖成穗及产量的影响. 农业工程学报, 2014, 30(10): 37–47. Song Y S, Zhang H C, Dai Q G, Yang D L, Guo B W, Zhu C C, Huo Z Y, Xu K, Wei H Y, Hu J M, Wu A G, Jiang X H. Effect of rice potted-seedlings per hole by mechanical transplanting on tillers emergence, panicles formation and yield., 2014, 30(10): 37–47 (in Chinese with English abstract).
[14] 雷小龙, 刘利, 刘波, 黄光忠, 马荣朝, 任万军. 杂交籼稻机械化种植的分蘖特性. 作物学报, 2014, 40: 1044–1055.Lei X L, Liu L, Liu B, Huang G Z, Ma R C, Ren W J. Tillering characteristics ofhybrid rice under mechanized planting., 2014, 40: 1044–1055 (in Chinese with English abstract).
[15] 许轲, 唐磊, 张洪程, 郭保卫, 霍中洋, 戴其根, 魏海燕, 韦还和. 不同机械直播方式对水稻分蘖特性及产量的影响. 农业工程学报, 2014, 30(13): 43–52. Xu K, Tang L, Zhang H C, Guo B W, Huo Z Y, Dai Q G, Wei H Y, Wei H H. Effect of different mechanical direct seeding methods on tiller characteristics and yield of rice., 2014, 30(13): 43–52 (in Chinese with English abstract).
[16] 熊瑞恒, 杭玉浩, 王强盛, 许国春, 刘欣, 武皞. 麦秸还田配施基蘖氮肥提高机插超级粳稻分蘖成穗及产量. 农业工程学报, 2015, 31(18): 136–146. Xiong R H, Hang Y H, Wang Q S, Xu G C, Liu X, Wu H. Wheat straw returned combined with nitrogen as base fertilizers and topdressing at tiller stage improving the tiller emergency, earbearing traits and yield for machine-transplanted superrice., 2015, 31(18): 136–146 (in Chinese with English abstract).
[17] 王萌萌, 杨沈斌, 江晓东, 王应平, 陈德, 黄维, 于庚康, 石春林. 光温要素对水稻群体茎蘖增长动态影响的分析及模拟. 作物学报, 2016, 42: 82–92.Wang M M, Yang S B, Jiang X D, Wang Y P, Chen D, Huang W, Yu G K, Shi C L. Analysis and simulation of impact of light and temperature on rice tillering., 2016, 42: 82–92 (in Chinese with English abstract).
[18] 孙成明, 庄恒扬, 杨连新, 杨洪建, 黄建晔, 董桂春, 朱建国, 王余龙. FACE水稻茎蘖动态模型. 应用生态学报, 2006, 17: 1448–1452.Sun C M, Zhuang H Y, Yang L X, Yang H J, Huang J Y, Dong G C, Zhu J G, Wang Y L. Dynamic model of rice tiller in FACE., 2006, 17: 1448–1452 (in Chinese with English abstract).
[19] Huang M, Yang C L, Ji Q M, Jiang L G, Tan J L, Li Y Q. Tillering responses of rice to plant density and nitrogen rate in a subtropical environment of southern China., 2013, 149: 187–192.
[20] Ao H J, Peng S B, Zou Y B, Tang Q Y, Visperas R M. Reduction of unproductive tillers did not increase the grain yield of irrigated rice., 2010, 116: 108–115.
[21] 吕伟生, 曾勇军, 石庆华, 潘晓华, 黄山, 商庆银, 谭雪明, 李木英, 胡水秀. 基于机插晚稻分蘖成穗特性获取基本苗定量参数. 农业工程学报, 2016, 32(1): 30–37.Lyu W S, Zeng Y J, Shi Q H, Pan X H, Huang S, Shang Q Y, Tan X M, Li M Y, Hu S X. Calculation of quantitative parameters of basic population of machine-transplanted late rice based on its tillering and panicle formation characteristics., 2016, 32(1): 30–37 (in Chinese with English abstract).
[22] 荆培培, 崔敏, 秦涛, 周在中, 戴其根. 土培条件下不同盐分梯度对水稻产量及其生理特性的影响. 中国稻米, 2017, 23(4): 26–33. Jing P P, Cui M, Qin T, Zhou Z Z, Dai Q G. Effects of different saline stress on yield and physiological properties of rice in soil culture., 2017, 23(4): 26–33 (in Chinese with English abstract).
[23] 李景蕻, 李刚华, 杨从党, 王绍华, 刘正辉, 王强盛, 丁艳锋. 增加土壤温度对高海拔生态区水稻分蘖成穗及产量形成的影响. 中国水稻科学, 2010, 24: 36–42.Li J H, Li G H, Yang C D, Wang S H, Liu Z H, Wang Q S, Ding Y F. Effects of temperature increase of soil on productive tiller percentage and yield of rice in high altitude ecological area., 2010, 24: 36–42 (in Chinese with English abstract).
[24] 钟旭华, 彭少兵, Sheehy J E, 刘鸿先. 水稻群体成穗率与干物质积累动态关系的模拟研究. 中国水稻科学, 2001, 15: 107–112.Zhong X H, Peng S B, Sheehy J E, Liu H X. Relationship between productive tiller percentage and biomass accumulation in rice (L.): a simulation approach., 2001, 15: 107–112 (in Chinese with English abstract).
[25] 袁奇, 于林惠, 石世杰, 邵建国, 丁艳锋. 机插秧每穴栽插苗数对水稻分蘖与成穗的影响. 农业工程学报, 2007, 23(10): 121–125.Yuan Q, Yu L H, Shi S J, Shao J G, Ding Y F. Effects of different tiller production planting seedlings per hill on outgrowth and quantities of machine-transplanted rice., 2007, 23(10): 121–125 (in Chinese with English abstract).
[26] 李杰, 张洪程, 龚金龙, 常勇, 吴桂成, 郭振华, 戴其根, 霍中洋, 许轲, 魏海燕. 稻麦两熟地区不同栽培方式超级稻分蘖特性及其与群体生产力的关系. 作物学报, 2011, 37: 309–320.Li J, Zhang H C, Gong J L, Chang Y, Wu G C, Guo Z H, Dai Q G, Huo Z Y, Xu K, Wei H Y. Tillering characteristics and its relationships with population productivity of super rice under different cultivation methods in rice–wheat cropping areas., 2011, 37: 309–320 (in Chinese with English abstract).
[27] Wang J J, Dai Q X, Shang J L, Jin X L, Sun Q, Zhou G S, Dai Q G. Field-scale rice yield estimation using sentinel-1A synthetic aperture radar (SAR) data in coastal saline region of Jiangsu province, China., 2019, 11: 2274.
[28] 张瑞珍, 邵玺文, 童淑媛, 汪恒武, 齐春燕, 孙长占. 盐碱胁迫对水稻源库与产量的影响. 中国水稻科学, 2006, 20: 116–118.Zhang R Z, Shao X W, Tong S Y, Wang H W, Qi C Y, Sun C Z. Effect of saline alkali stress on source-sink and yield of rice., 2006, 20: 116–118 (in Chinese with English abstract).
[29] 胡博文, 谷娇娇, 贾琰, 沙汉景, 张君颜, 黄书勤, 赵宏伟. 盐胁迫对寒地粳稻籽粒淀粉形成积累及产量的影响. 华北农学报, 2019, 34(1): 115–123.Hu B W, Gu J J, Jia Y, Sha H J, Zhang J Y, Huang S Q, Zhao H W. Effect of salt stress on starch formation and yield ofrice in cold-region., 2019, 34(1): 115–123 (in Chinese with English abstract).
[30] 隗溟, 李冬霞. 水稻主茎节位分蘖及生产力补偿能力. 生态学报, 2013, 33: 7098–7107.Wei M, Li D X. The compensation capacity of tillering and production of main stem nodes in rice., 2013, 33: 7098–7107 (in Chinese with English abstract).
[31] 黄丽芬, 陶晓婷, 高威, 王远玲, 庄恒扬. 江苏沿海地区减磷对机插常规粳稻产量形成及品质的影响. 中国水稻科学, 2014, 28: 632–638.Huang L F, Tao X T, Gao W, Wang Y L, Zhuang H Y. Effect of reduced phosphorus fertilizer application on yield formation and quality ofrice in Jiangsu coastal region., 2014, 28: 632–638 (in Chinese with English abstract).
[32] 张洪程, 龚金龙. 中国水稻种植机械化高产农艺研究现状及发展探讨. 中国农业科学, 2014, 47: 1273–1289.Zhang H C, Gong J L. Research status and development discussion on high-yielding agronomy of mechanized planting rice in China., 2014, 47: 1273–1289 (in Chinese with English abstract).
[33] 田蕾, 陈亚萍, 刘俊, 马晓刚, 王娜, 杨兵, 李莹, 郭海东, 李娟, 胡慧, 张银霞, 李培富. 粳稻种质资源芽期耐盐性综合评价与筛选. 中国水稻科学, 2017, 31: 631–642.Tian L, Chen Y P, Liu J, Ma X G, Wang N, Yang B, Li Y, Guo H D, Li J, Hu H, Zhang Y X, Li P F. Comprehensive evaluation and selection of rice () germplasm for saline tolerance at germination stage., 2017, 31: 631–642 (in Chinese with English abstract).
Tillering characteristics and its relationships with population productivity ofrice Nanjing 9108 under salinity stress
WEI Huan-He1, GE Jia-Lin1, ZHANG Xu-Bin1, MENG Tian-Yao2, LU Yu1, LI Xin-Yue1, TAO Yuan1, DING En-Hao1, CHEN Ying-Long1, and DAI Qi-Gen1,*
1Jiangsu Key Laboratory of Crop Genetics and Physiology / Jiangsu Key Laboratory of Crop Cultivation and Physiology / Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops / Research Institute of Rice Industrial Engineering Technology, Yangzhou University, Yangzhou 225009, Jiangsu, China;2Joint International Research Laboratory of Agriculture and Agro-product Safety, Ministry of Education / Institute of Agricultural Science and Technological Development, Yangzhou University, Yangzhou 225009, Jiangsu, China
This study was conducted to clarify the characteristics for occurrence of rice tillers, its dominant leaf positions, and panicle formation under salinity stress, and to provide a basis for the rational utilization and regulation of rice tillers for high-yield cultivation methods in tidal flat area. Conventionalrice Nanjing 9108 was used with three salinity treatments, namely, Control (CK, 0 salt concentration), Medium-salinity stress (MS, 0.15% salt concentration), and high-salinity stress (HS, 0.3% salt concentration) to compare the tillering and panicle formation characteristics and its contribution to population yield under salinity stress. As a result, the average yields (t hm–2) of CK, MS, and HS were 9.7, 7.4, and 4.2 across two years, respectively; the number of panicles, spikelets per panicle, percentage of filled grains, and 1000-grain weight in MS and HS treatments were significantly lower than those in CK. Compared with CK, the number of tillers at jointing, heading, and maturity and percentage of productive tillers in MS and HS treatments were decreased. For CK, the primary tillers emerged from the 3rd leaf to 7th leaf on the main stem, with higher tiller emerging rate and more panicles from the 4th leaf to 6th leaf among them, and secondary tillers mainly emerged and earbeared from 1/4 and 1/5 tiller position; as for MS and HS, tillers are mainly primary ones emerging from the 4th to 6th leaf as the dominant position for the tiller occurrence and earbearing. The mean value of panicle length, total grains, grain density, and number of grains and its total grains on the primary and secondary branches in MS and HS treatments were both lower than those in CK. Our results suggest that, salinity stress caused lower number of panicles per plant, inferior growth of individuals and populations, smaller panicle types, and finally lower grain yield.
salinity stress;rice; tillering characteristics; population productivity
10.3724/SP.J.1006.2020.02001
本研究由国家科技支撑计划项目(2015BAD01B03), 江苏省重点研发计划项目(BE2015337, BE2016370), 国家自然科学基金项目(31901448), 江苏省高等学校自然科学研究面上项目(19KJB210004)和江苏高校优势学科建设工程项目资助。
This study was supported by the National Key Technology Support Program of China (2015BAD01B03), the Key Research and Development Program of Jiangsu Province (BE2015337, BE2016370), the National Natural Science Foundation of China (31901448), the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (19KJB210004), and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
戴其根, E-mail: qgdai@yzu.edu.cn
E-mail: hhwei@yzu.edu.cn
2020-01-09;
2020-04-15;
2020-04-26.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20200426.1544.022.html
猜你喜欢
烟草科技(2022年9期)2022-09-24
作物学报(2022年3期)2022-01-22
山西农业科学(2021年4期)2021-04-19
江苏农业科学(2020年22期)2020-03-03
安徽农学通报(2019年11期)2019-07-12
农民致富之友(2019年9期)2019-05-22
农家科技(2018年2期)2018-05-05
新农业(2017年6期)2017-07-15
新农村(2015年10期)2015-09-28
现代农业科技(2015年1期)2015-04-02
