
三峡库区不同城镇化梯度下河流pCO2动态及驱动因素
2020-07-19 13:02易子涵李思悦李玉英
湖泊科学 2020年4期
易子涵,李思悦,唐 薇,李玉英
(1:中国科学院重庆绿色智能技术研究院,重庆 400714)
(2:中国科学院大学,北京 100049)
(3:南水北调中线水源区水安全河南省协同创新中心, 南阳师范学院农业工程学院,南阳 473061)
河流仅占地球表面的极小一部分, 却在陆地与海洋间碳循环的过程中扮演着重要的纽带作用. 河流输运陆地碳源到海洋生态系统时, 大部分的碳未能到达海洋, 流失的碳主要被固定、存储和以CO2形式释放[1]. 近年来, 城镇化进程使流域土地利用方式发生了显著改变, 明显改变了河流生态系统结构和功能, 进而影响河流内部碳的生物地球化学过程[2-5]. 河流被认为是大气重要的CO2源, 其对城镇化的响应对准确评估全球变化下内陆水体碳释放具有重要的科学意义, 但相关研究尤其缺乏.
CO2分压(pCO2)是控制水体向大气扩散或吸收CO2的重要因素之一, 当河流pCO2大于大气中的pCO2时, 河流为碳源. 河流pCO2变化规律及控制因素十分复杂, 它主要受河流内部碳酸盐体系以及外部的生物地球化学过程的影响[6-7]. 因此, 城镇化背景下人类对河流的外源输入增加及土地利用方式的改变会导致pCO2的显著变化. 国内外学者的研究表明内陆水体中的CO2通常处于过饱和状态. 例如, Raymond等在全球尺度上对6708条河流进行调查, 发现约95%的河流pCO2处于过饱和状态, 平均值为3100 μatm左右[8]. Li等对对湄公河的研究表明水体pCO2的平均值为1090 μatm[9]. Wu等发现三峡库区草堂河6月的表层水体pCO2低于大气[4], 水体呈碳汇特征, 但Yao等发现三峡库区干流秋季的pCO2为920~1140 μatm[10], 为大气CO2浓度的2~3倍. Li等在不同季节对三峡库区河流进行调查, 结果表明河流pCO2变化范围为50~6019 μatm, 秋季均值为1573 μatm, 夏季均值为1276 μatm[11]. Ni等对三峡库区大宁河的研究表明5月和8月pCO2均值分别为483和2018 μatm[12]. Wang等[3]对重庆城区范围的主要河流按土地利用组成进行分类调查, 结果表明河流pCO2随建设用地与耕地占比的上升有增加的趋势, 平均值达到3526 μatm, 远高于大部分自然河流.
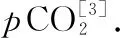
1 材料与方法
1.1 研究区域与样品采集
三峡库区东起湖北宜昌三斗坪三峡大坝坝址, 西至重庆市江津朱沱红花堡, 三峡水库全长663 km, 干流库段平均水面宽度1100 m, 淹没陆域面积约为580 km2[13]. 近年来, 库区范围内的城镇化进程逐年加剧. 建设用地的扩张以及人类活动强度的增加对河流生态系统造成了巨大的威胁. 本研究中的3条河流样点位置见图1. 桃花溪流域(29°51′11″~30°5′54″N, 107°4′42″~107°10′53″E)地处长寿区, 上游位于云台镇板桥湾, 下游处于长寿区市中心处, 河流整体受城镇化影响严重, 水质较差. 南河流域地处四川盆地东部平行岭谷丘陵区, 域内林木采伐过度, 植被较差, 水土流失严重. 汝溪河发源于万州区分水镇, 途经梁平区再穿过忠县汇入长江, 主河道长54.5 km, 常年水面宽度在100 m左右, 是忠县的第二大支流河, 下游位于汝溪镇内.
3条河流均位于三峡库区内, 各流域范围内土地利用组成不同, 呈现出不同的城镇化梯度. 其中桃花溪受到城镇化影响较为严重, 下游有大量生活垃圾及工业废水流入. 南河整体而言受到中等程度的城镇化影响. 汝溪河作为城镇化强度较低的河流进行对比, 以更加直观反映城镇化梯度对河流的影响.
本研究在桃花溪设置6个样点, 在南河设置7个样点, 在汝溪河设置4个样点. 为使得各河流能在不同城镇化梯度下进行纵向比较, 各河流样点基本均匀分布在河流的河源至河口处. 采样时间为2018年8月27日8月31日, 使用2.5 L采样器对水下0.1~0.2 m深的水样进行采集, 并加入浓硫酸抑制微生物活动, 水样于48 h内带回实验室冷藏, 于1周内完成实验分析.
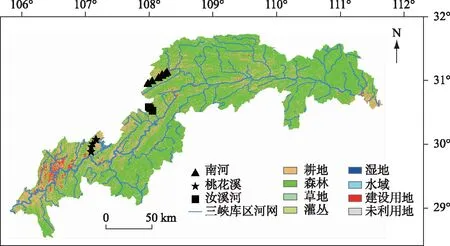
图1 三峡库区3条研究河流采样点及流域土地利用组成
1.2 实验分析与方法
1.2.1野外实验 现场使用Cyber Scan PCD 650多参数水质分析仪EutECh(美国)测定水温、pH、电导率(EC)等. 采样当天完成标准盐酸(0.0200 mol/L)滴定总碱度, 并完成水样的预处理工作. 当天用0.70 μm Whatman GF/F玻璃纤维膜过滤, 将滤液分装于聚乙烯瓶中, 不留空隙;用于测定溶解性有机碳(DOC)浓度. 另有水样采用0.45 μm Millipore硝酸纤维滤膜过滤, 用于测定溶解性总氮(DTN)、溶解性总磷(DTP)浓度.
1.2.2 室内实验 利用Multi N/C 2100S(德国耶拿)碳氮分析仪测定DOC浓度. 使用碱性过硫酸钾消解紫外分光光度法(HJ 636-2012)测定DTN, 钼酸铵分光光度法(GB 11893-1989)测定DTP[14];结合水温、pH及碱度计算pCO2(具体见1.2.4部分).
1.2.3 土地利用数据来源与数据处理 本研究中所有数据均录入SPSS和R进行统计分析与绘图, 采用One-way ANOVA来判别不同数据组的差异性, 采用Spearman非参数相关性分析方法阐释数据间变化的统计相关性. 土地利用组成数据来自清华大学地球系统科学系宫鹏最新研究成果——10 m分辨率的全球土地覆盖产品(FROM-GLC10)[15]. 研究平台以ArcGIS 10.3为主, 其他辅助资料及平台包括有全国数字高层(DEM)图、三峡库区行政区域图以及Google Earth等.
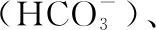
(1)
pK0=-7×10-5T2+0.16T+1.11
(2)
pK1=1.1×10-4T2-0.012T+6.58
(3)
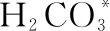
2 结果与分析
2.1 研究区域土地利用组成
代表高人类活动强度的建设用地对河流生态系统的影响最为强烈, 建设用地在土地利用栅格图中所占比例越大即代表城镇化强度越高[17]. 桃花溪流域建设用地占比最大, 为12.43%;南河次之, 建设用地占比为3.36%;汝溪河最低, 建设用地所占比例为0.6%. 城镇化强度高低顺序为:桃花溪>南河>汝溪河, 此外, 每条河流从上游至下游即沿水流方向城镇化强度增加.
2.2 河流pCO2变化规律
本研究中17个样点的pH值在7.56~9.48之间, 平均值为8.23±0.7, 整体表现为弱碱性, 不同河流的pH值无显著差异; 表层水温在30.8~37.4℃之间;碱度变化范围为1320~4320 μeq/L, 均值为409.4±856.2 μeq/L;EC变化范围为213.9~669.6 μS/cm, 均值为413.4±139.3 μS/cm. 图2表明, 沿水流方向, 桃花溪pH有增高的趋势, 汝溪河和南河则有降低的趋势;桃花溪、南河碱度有增加趋势;南河(除有异常值的Site 5外)、汝溪河pCO2呈现出明显递增的趋势, 而桃花溪则相反. 桃花溪的Site 1样点出现了高达12067 μatm的异常值, 这可能是由于该点位河宽窄且河水很浅, 河底有大量生活垃圾使得河流原位呼吸加强, 从而导致了该点位pCO2的极端值. 图3表明, Alk空间格局和城镇化梯度一致, 且桃花溪Alk显著高于其他2条河流(P<0.001), EC在桃花溪中显著升高(P<0.01). 南河、汝溪河、桃花溪pCO2平均值分别为2006±3546、1790±1210、4094±4218 μatm. 3条河流pCO2无显著性差异, 但各河流pCO2变化规律与城镇化梯度一致即城镇化强,度越高的河流具有更高的pCO2.
2.3 河流有机质及营养盐特征
河流DOC浓度范围为3.76~8.1 mg/L, 均值为5.37±1.44 mg/L;DTN浓度在0.037~2.99 mg/L范围内变化, 均值为0.73±0.81 mg/L;DTP浓度在0.08~0.53 mg/L之间变化, 均值为0.26±0.12 mg/L. 图2表明, 沿水流方向, 桃花溪、南河DOC浓度有增高趋势. 图3表明, DTP、DTN、DOC浓度平均值均在桃花溪最大, 且桃花溪DTN、DOC浓度均显著大于其他2条河流, DOC和营养元素(DTN、DTP)的变化规律与河流城镇化梯度一致.
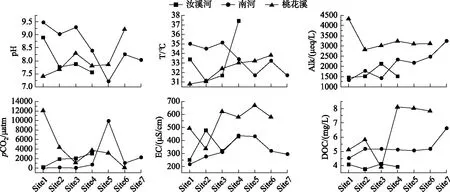
图2 河流不同站点理化指标及pCO2格局(从Site1到Site7为水流方向)
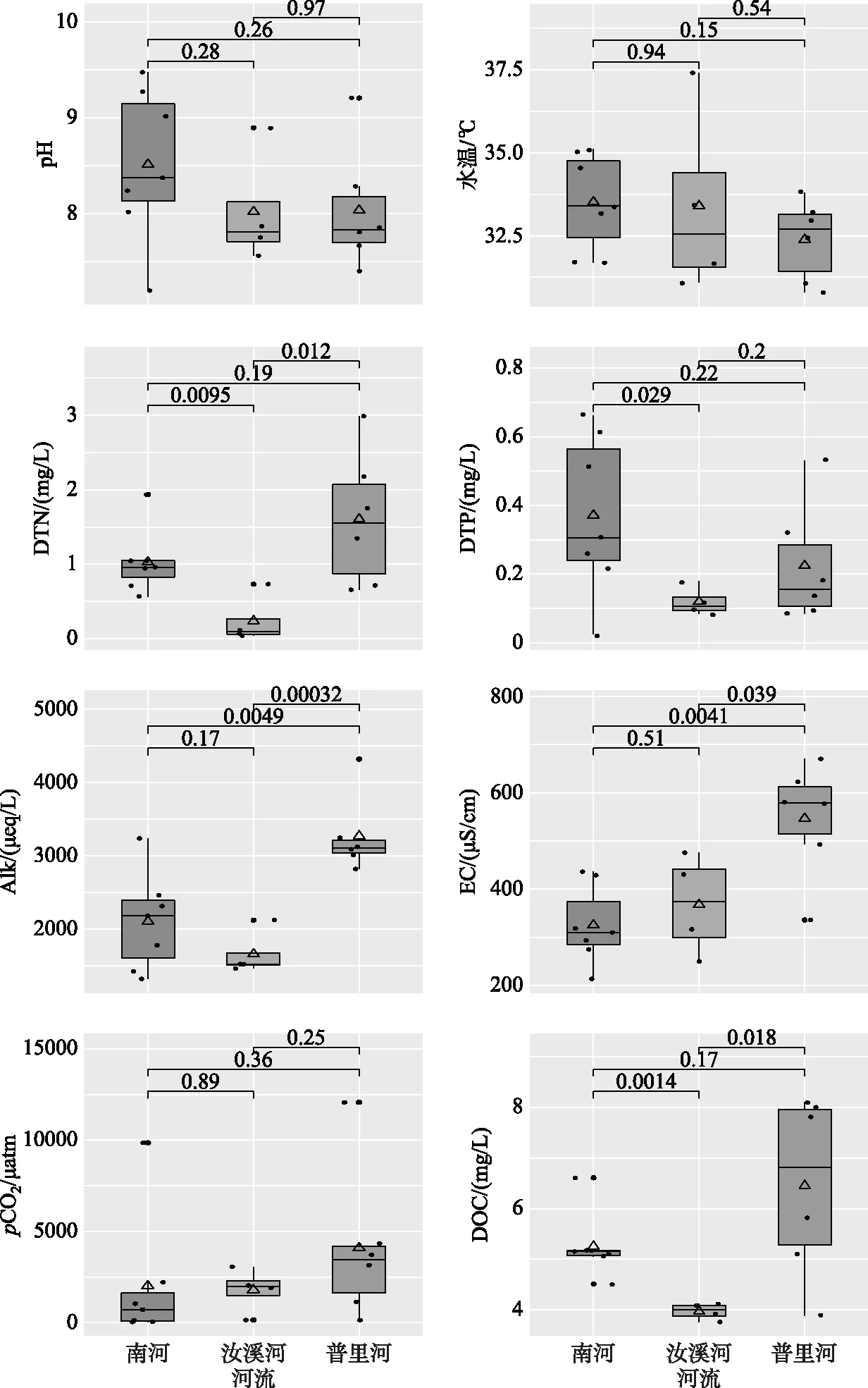
图3 河流水质指标及pCO2均值格局(箱体间的数值代表不同河流显著性差异水平, 三角形代表平均值, 横线代表中位数;从上到下分别为最大值、上四分位数、下四分位数、最小值)
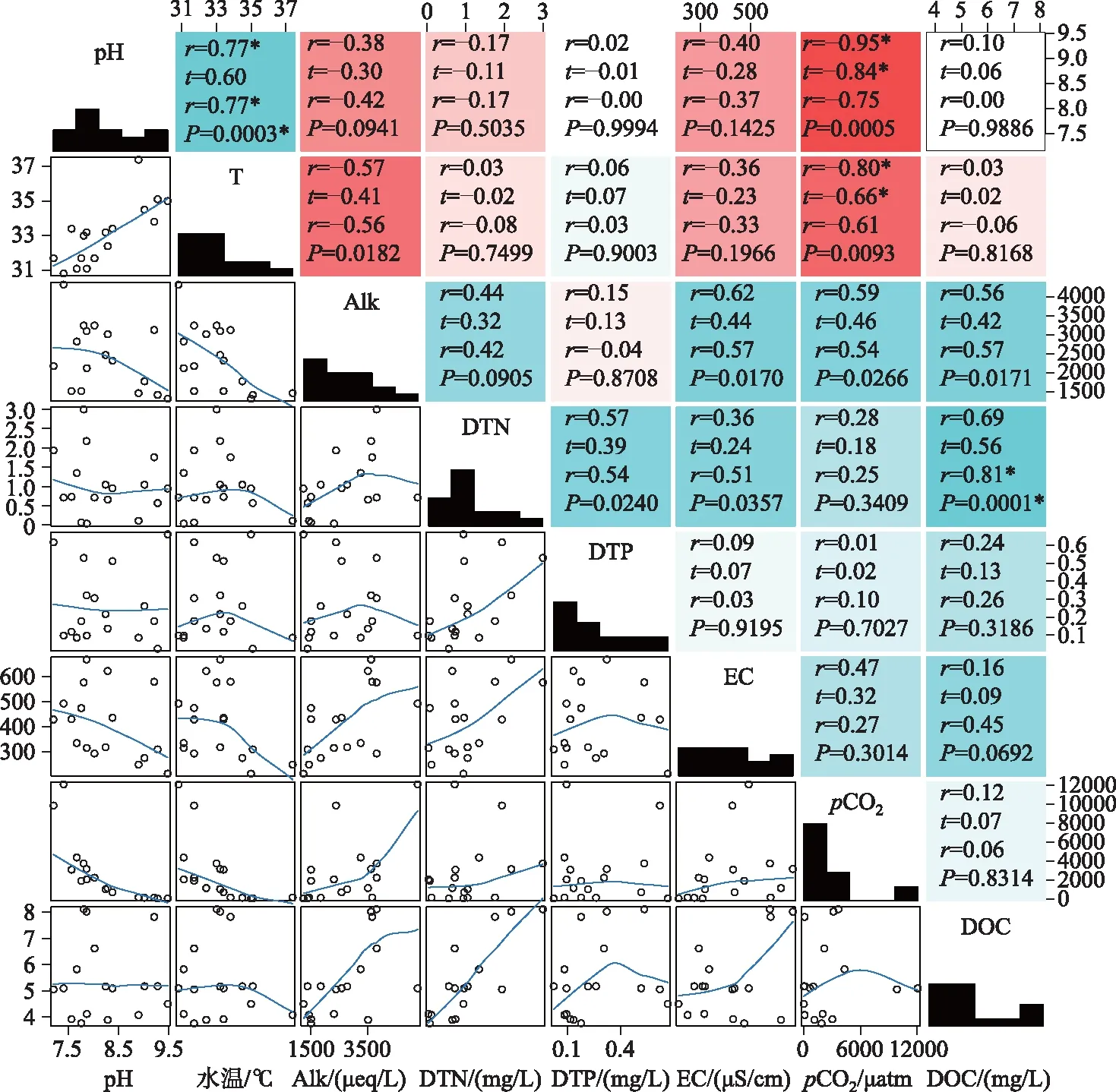
图4 环境因子相关关系矩阵(空白处表示无显著相关性, *、**和***代表的显著性差异水平分别为P<0.05、P<0.01和P<0.001;右上部分数字代表相关系数;左下部分为两因子间的散点图;对角线为各因子的频数统计图)
3 讨论
3.1 河流水环境对pCO2的影响
pH、温度与pCO2呈极显著负相关(图4), 此结果与国内外学者的研究一致[18-21]. pH值和水体的碳酸盐动态平衡密切相关, 游离态CO2增加会导致pH降低. 水温对pCO2影响机制较为复杂, 通常情况下, 水温通过影响气体分子在水中的溶解度、微生物活性、水生植物光合、呼吸作用和气泡的形成影响CO2的产生及消耗过程[22-24], 温度较高时, 一方面水生植物光合作用增强, 大量消耗水中CO2;另一方面水生生物的呼吸作用、微生物矿化降解DOC都有利于pCO2增加. 夏季的高温与长时间的光照条件使水生植物的光合作用占主导地位从而引起pCO2的下降. 碱度是影响水体pCO2的又一重要因素, 碱度的大小代表着水体中碳酸盐浓度, 当碳酸盐浓度较高时会促使CO2与水的反应逆向进行从而增加pCO2.
养分(DTN、DTP浓度)对水体pCO2的浓度变化至关重要,它们一方面促进水体中微生物的矿化作用使水体中CO2增多,另一方面促进水生植物的光合作用消耗CO2,本研究的结果表明pCO2与DTN和DTP浓度无显著相关关系,这可能是因为夏季光照充足且水温较高极大促进了水生植物的光合作用使其与矿化和呼吸作用所产生的CO2相互抵消. DOC大部分来自与土壤有机质(DOM)的分解产物或者人类生产活动中所产生的有机废物,另一部分来自于浮游生物的代谢过程[25],是河流水体中CO2的主要碳源之一. 本研究发现pCO2与DOC浓度无明显的相关性,主要归因于人为活动的影响弱化了其相关关系.
3.2 城镇化梯度对河流理化性质及pCO2的影响
河流中N、P形态及浓度受到人类生产生活活动和其他环境因素的影响. 耕地和建设用地较其他土地利用方式而言更易产生非点源污染. 耕地区域,农药、无机肥料和有机肥的施用是提高当地作物产量的主要因素. 传统的农业活动增加了土壤侵蚀的风险,此外,如过度施肥能导致非点源污染通过地表径流进入地表水. 在旱季,城市土地与许多水质参数呈正相关(例如DTP、氨氮). 降水期间,建设用地上的非点源污染(如有机化合物、重金属和多种来源的养分)从不透水表面流入河流进而影响河流水体的理化性质[26]. 本研究中,有61%的样点N∶P化学计量比低于16∶1[27],这说明大部分水体的营养盐限制元素为N元素. 对不同河流的DTN和DTP进行比较,表明汝溪河的DTN和DTP均处于最低的水平,其DTN达到Ⅱ类地表水标准,DTP为Ⅲ类地表水标准;南河DTN为Ⅳ类地表水标准,DTP为Ⅲ类;桃花溪的DTN浓度远高于其他2条河流,DTN为Ⅴ类地表水标准,DTP为Ⅳ类,此结果也与各河流的城镇化梯度趋势一致. 此外,受城镇化污染输入的影响,桃花溪的EC和DOC浓度均值也显著大于其他2条河流.
通过对三峡库区城镇化梯度下典型河流的研究发现城镇化强度较高的河流pCO2更高,其中汝溪河和南河的pCO2沿水流方向也呈升高趋势,表明外源输入会导致河流原位呼吸增强进而使河流pCO2增加. 这一结果与吴学谦等夏季在三峡库区范围内草堂河的研究结果一致[4]. 虽然pCO2在不同河流间与同一河流上下游的不同站点均对城镇化梯度产生响应,但本研究仅以土地利用组成图定性描述了城镇化梯度,进一步的研究将会量化城镇化梯度以期更准确地揭示pCO2与城镇化梯度的定量关系;此外,城镇化梯度对pCO2的影响机制及关键生物地球化学过程极其复杂,其作用机理以及与气象、水文地质等条件交叉产生的影响仍需深入的研究.
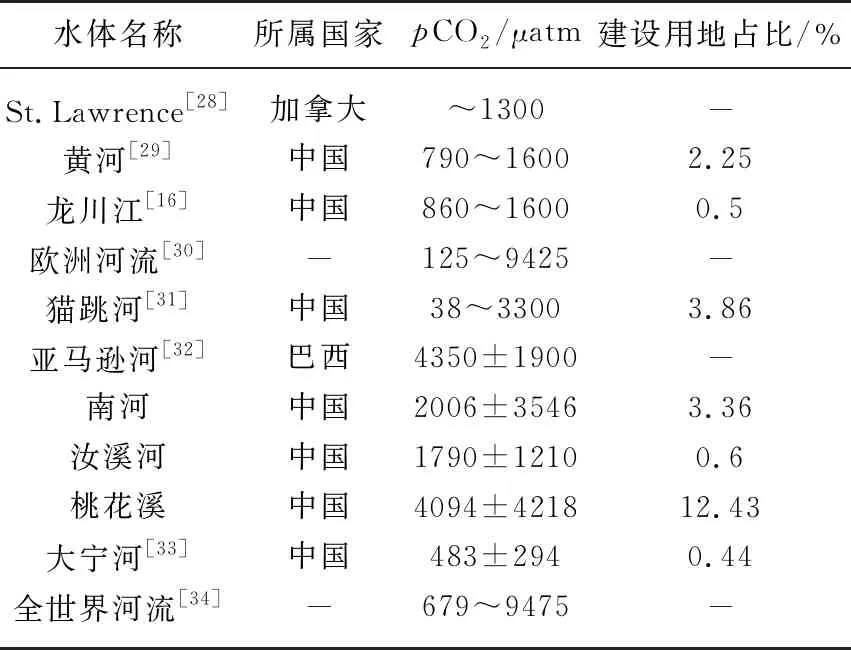
表1 世界河流水体pCO2比较*
3.3 与其他河流pCO2比较
表1表明全球大部分河流水体中的pCO2大于大气中的CO2浓度,总体表现为碳源. 一般地,热带河流、亚热带河流、温带河流的pCO2呈现依次递减的趋势. 我们的研究表明在一个区域内,城镇化可以明显增加河流pCO2,主要归因于城镇化进程中人类生产活动过程中向水体输入的营养元素、有机质等增强了河流的原位呼吸. 此外,本研究中的3条河流位于亚热带区域,较同区域的其他河流的pCO2高. 但在大尺度上,城镇化占比和河流pCO2并未表现出明显的正相关关系,主要是由于河流pCO2的影响因素极其复杂.
4 结论
1) 三峡库区典型城镇化河流水体中的pCO2高于大气中的CO2浓度,呈现CO2源.
2)研究区域内,城镇化强度越高的河流具有高的营养盐、有机质浓度及pCO2,表明人类活动与土地利用方式的改变极大地改变了河流生态系统的碳循环过程.
3)南河和汝溪河的pCO2沿上游至下游显著升高,主要归因于下游高的营养物质及有机碳的输入增加了河流的原位呼吸. 研究不同城镇化梯度下pCO2的变化规律对城镇化进程中的河流碳循环尤其是CO2释放的评估具有突出的科学意义.
致谢:感谢中国科学院重庆绿色智能技术研究院生态水文研究中心的各位老师在本研究样品采集和分析过程中给予的支持和帮助.
猜你喜欢
黄河之声(2022年1期)2022-11-22
农村青少年科学探究(2019年12期)2019-03-28
作文与考试·初中版(2017年20期)2017-07-08
中华诗词(2016年10期)2017-02-22
爱你(2016年11期)2016-11-26
创新作文(1-2年级)(2016年4期)2016-05-14
故事会(2016年6期)2016-03-23
小猕猴智力画刊(2013年1期)2013-03-15
中学生物学(2008年8期)2008-10-27
雕塑(1995年4期)1995-07-12
