
河南地方猪mtDNA D-loop序列遗传多样性及系统进化分析
2020-07-09 10:51李新建薛亚辉王明宇段栋栋刘贤乔瑞敏韩雪蕾王克君李秀领
河南农业大学学报 2020年3期
李新建,薛亚辉,王明宇,段栋栋,刘贤,乔瑞敏,韩雪蕾,王克君,李秀领
(1.河南农业大学牧医工程学院,河南 郑州 450002; 2.河南省畜牧总站,河南 郑州 450002)
哺乳动物线粒体DNA(Mitochondrial DNA,mtDNA)是目前已知的唯一核外遗传物质,具有相对分子质量小、进化速度快、结构简单、呈母系遗传等特点。其中,D-loop区是线粒体DNA的非编码区,也称控制区,负责调控线粒体DNA的转录和复制,是线粒体DNA中序列和长度变异程度最高的地区,进化速度较其他区域高5~10倍,现已被广泛应用于哺乳动物群体遗传多样性、物种亲缘关系和系统发生等方面的研究。王继英等[1]研究了山东各猪种的线粒体DNA D-loop区,发现山东地方黑猪血统单一、遗传关系较复杂。李正杰等[2]研究了藏猪的线粒体DNA D-loop区的遗传多样性,发现诺尔盖地区藏猪遗传多样性较高、受人工选择强度低。赵忠海等[3]对贵州地方猪种的线粒体DNA D-loop区分析后发现宗地花猪的遗传多样性最为丰富。南阳黑猪、确山黑猪、淮南猪、豫西黑猪是河南省优良地方猪品种,具有繁殖力强、肉品质高、适应性强等特点[4]。20世纪80年代以来,随着社会经济的发展,消费者对高瘦肉率猪肉的需求不断提升,致使中国引进了生长迅速、瘦肉率较高的西方猪种,这直接导致了河南地方猪数量的急剧下降,使许多优秀基因在尚未研究其价值之前从猪基因库消失。因此,研究河南省地方猪种的遗传多样性和进化关系,对于正确认识和评价地方猪种质资源及科学合理地利用地方猪资源具有重要意义。但目前对河南地方猪种的研究报道还较少。本研究测定分析了河南各地方猪种mtDNA D-loop区序列,并结合GenBank数据库中的序列,在分子水平探讨了其遗传资源多样性及进化关系,为地方猪种的保种和利用提供依据。
1 材料与方法
1.1 样品采集及保存
河南及邻省6个猪种(南阳黑猪、确山黑猪、豫西黑猪、淮南猪、莱芜猪、二花脸猪)和野猪的样品采集信息见表1。采集到的耳组织样品,置于1 mL 75%乙醇中保存。3个国外猪种(杜洛克猪、约克夏猪、长白猪)的mtDNA D-loop序列从GenBank下载,序列号为杜洛克猪:AM040623-AM04632、EU660148-EU660151、GQ141897、KF767444,约克夏猪:AM040633-AM040638、GQ141893、GQ169777,长白猪:AM040613-AM040622、GQ141895、GQ169780,共计38条序列。
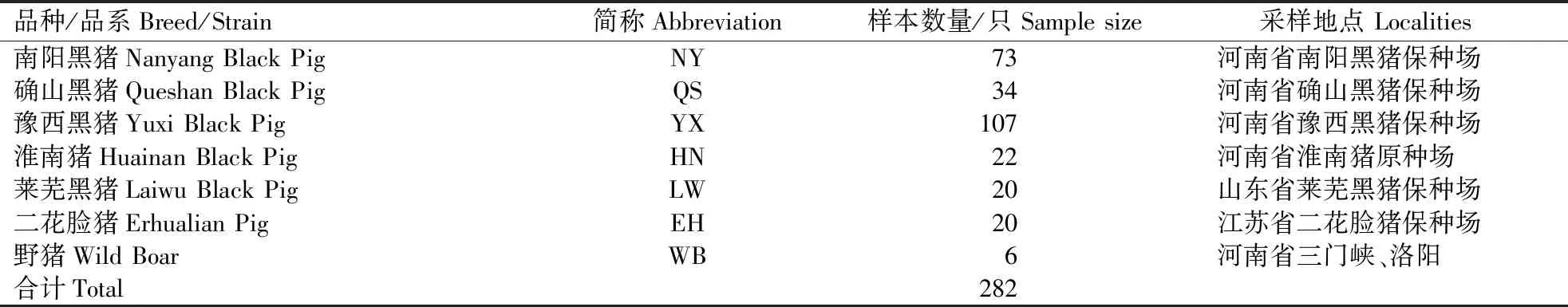
表1 样品信息Table 1 Sample information
1.2 基因组DNA的提取及检测
采用组织基因组DNA提取试剂盒(DP304,天根生化科技有限公司)提取DNA。利用微量核酸定量仪(Nanodrop2000)和1%琼脂糖凝胶电泳检测DNA的浓度和质量。
1.3 PCR扩增及测序
利用Primer5.0根据NCBI上长白猪参考序列(NC_000845)设计引物,上游引物为:5′-AGGAGACTAACTCCGCCAT-3′,下游引物为:5′-CGCGGATACTTGCATGTGT-3′,引物由上海生工生物有限公司合成。PCR反应体系(25μL):包括40 ngDNA、12.5 μL 2×ES Taq MasterMix(康为世纪CW2214M)、10 pmol正反向引物、8.5 μL水。扩增条件为:预变性94℃,5 min;变性94℃,30s;退火51.3℃,30 s;延伸72℃,50 s,25个循环,最后延伸72℃,10 min。PCR产物采用1%琼脂糖凝胶电泳检测PCR产物,检测合格后进行Sanger双向测序。
1.4 序列比对及数据分析
利用Mega7.0软件进行序列的核对编辑和拼接及统计各序列碱基的组成,采用邻接法(Neighbor-Joining,NJ法),Kimura 2-parameter模型,重复抽样1000次构建单倍型NJ系统发育树。使用DNAsp6.12软件统计单倍型数量、单倍型多样度、核苷酸多样度、平均核苷酸差异数,计算歧点分布。使用Popart1.7软件绘制单倍型网络图。
2 结果与分析
2.1 mtDNA D-loop区序列分析
用DNAsp 6.12软件统计所测猪种的D-loop区序列的碱基组成,结果见表2。由表2可知,T、C、A、G四种碱基的平均比例分别为24.92%、26.42%、32.57%、16.03%。A+T的含量为57.49%,大于G+C的含量42.45%,说明所测猪D-loop区为A-T富集区。在本研究中,河南4个猪种mtDNA D-loop中串联重复单位均为GCGTACACGT,在不同个体中串联重复次数的不同是造成D-loop序列长度差异的主要原因。
2.2 mtDNA遗传多样度分析
用DNAsp 6.0软件统计所测猪种的D-loop区序列的遗传多样性参数,结果见表3。由表3可知,所测得河南3个猪群体(南阳黑猪、确山黑猪、豫西黑猪)的单倍型多样度(Hd)均大于0.7,核苷酸多样度(Pi)均大于0.003,平均核苷酸差异数(K)均大于2.5,而淮南猪的单倍型多样度、核苷酸多样度、平均核苷酸差异数分别只有0.788、0.001 56、1.818,相对较低,表明淮南猪群体遗传差异相对贫乏。莱芜猪与二花脸猪的群体遗传变异较丰富。对6个群体的mtDNA D-loop区序列用Tajima’s D值进行中性检验,经检验均不显著(P>0.05),符合中性突变。
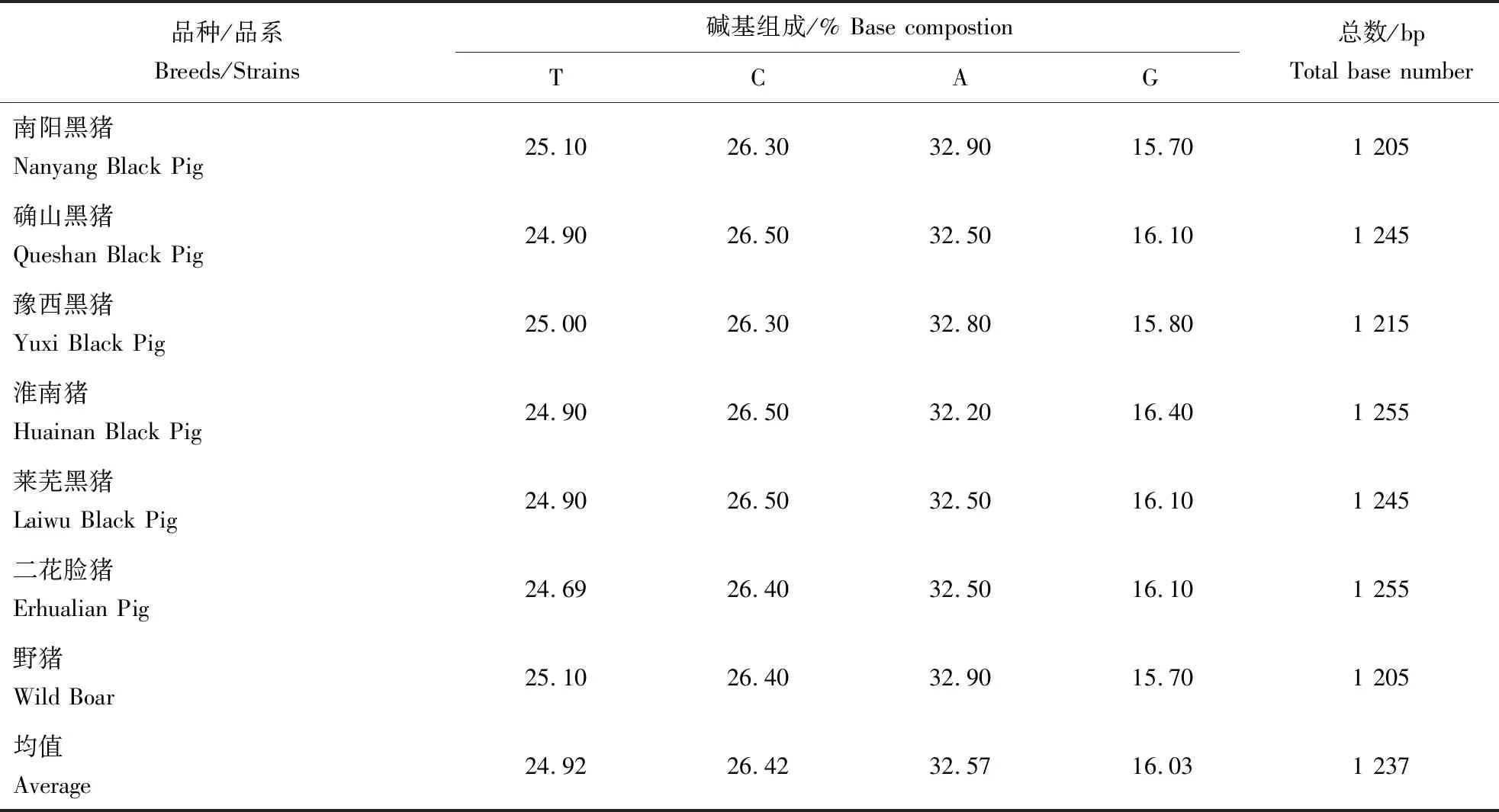
表2 不同猪种mtDNA D-loop序列碱基组成Table 2 Base composition of mtDNA D-loop sequences in different pig breeds
2.3 mtDNA D-loop区序列变异分析
将本研究测定的豫西黑猪、南阳黑猪、确山黑猪、莱芜猪、淮南猪、二花脸猪、野猪282条序列,结合从GenBank下载的3个国外猪种38条序列共计320条序列进行了比对,共发现37个多态位点。以单倍型H1为标准进行了比较分析,界定了42种单倍型,群体间共享单倍型15个、某个群体所独有单倍型27个(南阳黑猪5个、豫西黑猪10个、确山黑猪4个、淮南猪2个、莱芜猪3个、二花脸猪1个、野猪2个),各单倍型变异位点见表4。

表3 不同猪种mtDNA D-loop序列多态性、单倍型及其多样性Table 3 The polymorphism,haplotypes and genetic diversity of mtDNA D-loop sequence in different pig breeds
2.4 mtDNA D-loop系统发育分析
以非洲疣猪的mtDNA D-loop序列(GenBank登录号:DQ409321)作为外群构建河南4个地方猪种以及莱芜猪、二花脸猪和3个国外猪种的mtDNA D-loop序列单倍型NJ系统发育树,结果见图1。由图1可知,用于分析的42个单倍型在NJ树上明显分为2个独立的支系。一支包含了所有地方猪和部分大约克夏猪,另一支则全部是3个国外猪种的单倍型,表明河南4个地方猪种间的亲缘关系较近,而与国外猪种间亲缘关系相距较远,河南4个地方猪种与3个国外猪种有着两个明显不同的起源。豫西黑猪、南阳黑猪、确山黑猪、淮南猪、莱芜猪及二花脸猪的单倍型在整个系统进化树中分布较为紧密,且具有多个共享单倍型,表明这几个群体彼此之间可能有基因交流。国外猪种大约克夏猪有一个单倍型与地方猪聚在一起,表明大约克夏猪的母源血统部分来自于中国地方猪种。再结合已登录GenBank的中国其他地方猪16条序列,重新绘制单倍型NJ系统发育树,结果见图2。由图2可知,其拓扑结构与图1类似,进化树主干明显分为2个支系。
2.5 单倍型网络分析
根据DNAsp6.12软件统计分析并输出的单倍型结果,利用Popart1.7软件构建了河南和邻省6种地方猪mtDNA D-loop单倍型网络图,发现网络分析结果也明显分为2支(图3)。每个圆代表1种单倍型,圆的面积大小表明其频率的高低,圆中不同的颜色代表不同类型猪种。由图3可知,单倍型H12、H13、H18中的猪品种数较多,呈发散状,表明群体可能发生过群体扩张。单倍型H9、H12、H13、H17、H18、H20、H21、H23、H28为不同品种间共享单倍型。
2.6 种群历史动态分析
使用DNAsp6.12的Population size down模型对河南4个地方品种单倍型进行歧点分布分析,结果如图4。由图4可知,歧点分布实际值呈多峰分布,与假定期望模型不符,且中性检验值Tajima’s D为-1.877 21,Fu’s Fs为-26.018,经检验差异显著(P<0.05),表明河南地方猪群体曾经历过群体扩张事件。结合表3可知,各群体内Tajima’s D检验不显著,河南地方猪累加的突变使中性检验结果显著。





3 讨论与结论
3.1 mtDNA D-loop遗传多样性分析
衡量一个品种(群体)mt DNA变异程度的两个重要指标是单倍型多样度(Hd)和核苷酸多样度(Pi),单倍型多样度是指样本中随机抽取到2个不同单倍型的频率、核苷酸多样度是指给定群体内随机选取的mtDNA 序列间的平均每个位点的核苷酸差异数目[5]。Hd和Pi值越大,群体的多态程度越高,其遗传多样性越丰富[6]。据统计同种动物的个体之间的平均核苷酸顺序奇异值在0.3%~4.0%,有时甚至可达10%[7]。
本研究的河南4个地方猪群体中单倍型多样度和核苷酸多样度分别为0.788~0.854、0.001 56~0.004 55,4个猪种的Hd值分别高于莱芜猪(0.589)、陆川猪(0.640)、东乡花猪(0.768)、藏猪(0.705)、滇南小耳猪(0.759),低于二花脸猪(0.947)、桂中花猪(0.942)、合作猪(0.883),与保山猪(0.828)、五指山猪(0.838)、巴马香猪(0.875)处于同等水平;但4个猪种中除淮南猪核苷酸多样度(Pi)值较低,为0.001 56,其余3个猪种Pi值分别高于莱芜猪(0.002 46)、藏猪(0.002 83)、低于东乡花猪(0.005 9)、陆川猪(0.007 4)、滇南小耳猪(0.005 21)、合作猪(0.005 49)、与二花脸猪(0.003 73)、桂中花猪(0.003 43)、巴马香猪(0.003 37)、保山猪(0.003 71)、五指山猪(0.003 34)、巴马香猪(0.003 37)处于同等水平,表明河南地方猪种遗传多样性相对丰富[8-12]。
从河南地方猪种单倍型分布来看,4个群体的共享单倍型H12、H13、H18频率分别为45、41、47,说明这3种单倍型应该是河南地方猪共同的祖先单倍型。结合单倍型网络图分析显示河南地方猪种至少存在3个不同母系驯化事件[13]。
3.2 群体系统发育分析
张冬杰等[14]对中国43个地方猪品种和3个国外猪种基于mtDNA D-loop序列NJ分子进化树显示,中国地方猪和国外品种处在2个不同分支上。黄树文等[15]构建了基于遗传距离的广东5个地方猪种和国外4个猪种NJ进化树,发现广东省地方猪与西方猪种遗传距离较远。本研究结果表明,用于分析的7个群体在NJ树上明显聚为2个独立支系:一支包含了所有河南及邻省地方猪群体、野猪群体和部分大约克夏猪的单倍型,另一支则全部是3个国外猪种的单倍型。这与以上研究结果基本一致,同时也支持多数学者认同的现代家猪起源于欧亚两大母系的二元论观点[16-18]。然而,LARSON等[19]通过分析全世界范围内686个家猪和野猪个体mtDNA D-loop序列后认为家猪的起源在欧亚大陆有多个独立的驯化中心。LUETKEMEIER等[20]通过亚欧家猪和野猪mt DNA基因组分析得出中国猪有多个驯化中心。王志等[21]通过对黄河流域古代猪mtDNA D-loop序列的分析发现,黄河流域曾是中国家猪驯化中心之一。但YU等[22]在利用mtDNA D-loop区构建系统进化树分析中国地方猪起源时,发现中国家猪可能起源于长江流域和华南地区的野猪。河南地方猪具体起源何处,需要进一步的研究与证实。
本研究发现,约克夏猪部分单倍型与河南地方猪聚为一类,这与张冬杰等[14]研究发现约克夏猪部分单倍型与梅山猪聚为一类,张冰等[23]研究发现约克夏猪部分单倍型与广西地方猪聚为一类,NI等[18]研究发现约克夏猪和中国地方猪聚为一类,而非欧洲猪。许多研究[14,18,23]结果都表明约克夏猪部分母源血统来自中国地方猪,据此推测在约克夏猪培育过程中,可能引入了中国猪种的血统。
综上所述,河南省地方猪4个品种共236条序列,定义了32种单倍型,单倍型多样度和核苷酸多样度分别为0.788~0.854、0.001 56~0.004 55,表明遗传多样性较丰富。河南地方猪与西方猪种遗传差异较大,与西方商业猪种明显聚为2个独立分支,表明mtDNA D-loop序列可作为研究种内系统发育、起源进化的分子标记。歧点分布显示,整个河南地方猪群体曾经历过群体扩张事件。
猜你喜欢
猪业科学(2022年10期)2022-11-03
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
猪业科学(2022年1期)2022-03-24
猪业科学(2021年6期)2021-08-12
猪业科学(2021年3期)2021-05-21
读友·少年文学(清雅版)(2019年1期)2019-05-09
娃娃画报(2014年9期)2014-10-15
小朋友·快乐手工(2014年1期)2014-02-25
