
太行山南麓优势树种不同坡向下的叶功能及土壤养分差异
2020-07-09 10:51:12杨子范世献周芮宸张志铭赵勇
河南农业大学学报 2020年3期
杨子,范世献,周芮宸,张志铭,赵勇
(河南农业大学林学院,河南 郑州 450002)
Mountains
植物在生长发育过程中,与外部生长环境相互依存,从而逐渐形成适应外部环境变化的不同形态和生理结构,这种差异主要体现在植物的叶片、根系、种子等性状上,这种能响应外部环境因子变化的植物性状被称为植物功能性状[1]。植物的叶片作为其自身生长显露在外部环境中的表面积/体积比最大的器官,是植物与外界环境进行光合作用、气体交换及蒸腾的最重要功能单位[2]。而叶功能性状作为植物的主要特性之一,更是通过叶片这个重要的植物器官进行表征,研究叶功能性状与植物的生长发育和资源利用率是有紧密联系的[3-4]。植物的叶功能性状主要包括生理性状(光合作用特征:净光合作用速率、气孔导度、胞间CO2浓度)和叶面积分配性状(叶片大小(leaf size,LS)、叶厚度(leaf thickness,LT)、比叶面积(specific leaf area,SLA)和(叶干物质含量(leaf dry matter content,LDMC))[5]。已有研究表明,温度、降水、光照等会因坡向变化使其产生再分配,坡向变化会导致气候环境的空间异质性,从而影响到植物功能性状的变化[6-8]。刘雯霞等[9]研究发现,在阴坡上比叶面积与土壤含水量呈显著正相关,在半阴坡上与土壤pH值呈显著负相关,在阳坡上则与土壤有机质含量呈显著正相关。盘远方等[10]通过研究桂林岩溶石山灌丛植物的叶功能性状随坡向梯度变化规律发现:在阴坡上,对比叶面积影响显著的环境因子是土壤有效氮含量;阳坡上则是土壤含水量和土壤pH值。由此可知,研究不同植物物种间植物功能性状和土壤环境因子差异性,可以更好地揭示植物对环境的适应策略,从而为地形复杂地区的生态恢复提供依据。现阶段关于太行山南麓地区植物群落研究主要集中在环境因子与植物生长和发育方面。而植物自身对环境的适应导致自身结构生长发育所体现出来的物种间的差异性,具有重要研究价值。坡向作为植物外部环境生长的重要地形因子之一,对于植物生长发育所需的光照和雨水的再分配起关键作用,能够改善和营造局部小气候[11],植物对坡向的适应性可以更好地揭示植物的生长规律[12]。关于这方面的研究还很缺乏。太行山南麓属于典型的干旱半干旱区气候,当地土层浅薄贫瘠、天然林覆盖率较低、部分地区是裸露地,植被森林以人工种植为主。同时,常年降水量较少,导致地处干旱半干旱的太行山南麓地区面临严峻的水资源缺乏和养分流失等问题[13],使当地的生态系统逐渐变得更加脆弱。因此,本研究选取干旱半干旱的太行山南麓地区的优势树种酸枣、荆条、侧柏、栓皮栎作为研究对象,通过对不同坡向下植物树种间植物功能性状及土壤养分差异性的比较,探究不同树种植物对坡向的适应性,以期为太行地区生态修复和植被重建等工作提供参考。
1 材料与方法
1.1 研究区概况
本研究试验地位于河南省济源市大河沟林场(E112°28′-112°30′,N35°01′-35°03′,海拔320~400 m),该地区属于暖温带大陆性季风气候,年均气温14.33 ℃,年均日照率为54%,年际间变幅为44%,当地山区土壤主要为花岗片麻岩和石灰岩风化后形成的褐土,土层平均厚度为15~140 cm,土壤pH值7.0~7.5[14]。当地主要群落类型以乔木、灌木为主,其中灌木主要有荆条(Vitexnegundo)、酸枣(Ziziphusjujube)和胡枝子(Lespedezabicolor)等;乔木主要有栓皮栎(Quercusvariabilis)、侧柏(Platycladusorientalis)和刺槐(Robiniapseudoacacia)等[13]。该地区的优势树种主要是栓皮栎、侧柏、荆条和酸枣等,4种优势树种植被的大概比例为:1∶1.1∶6∶6.4。
1.2 试验设计与方法
1.2.1 试验设计 于2019年5月,选择太行山南麓地区的其中2个独立山丘作为试验地,选取山丘上生长成熟、长势良好的荆条、酸枣、栓皮栎、侧柏4种主要优势树种作为研究对象,在每个山丘的东、南、西、北坡4个方向以“主要字型设置10 m×10 m设置势样方,每个树种在每个坡向重复6次样方试验。对样地内每个个体进行群落学特征调查,计算郁闭度、平均密度、平均冠幅等(如表1所示)[14]。同时,测定并记录各树种株高及根部土壤温度和土壤含水量。
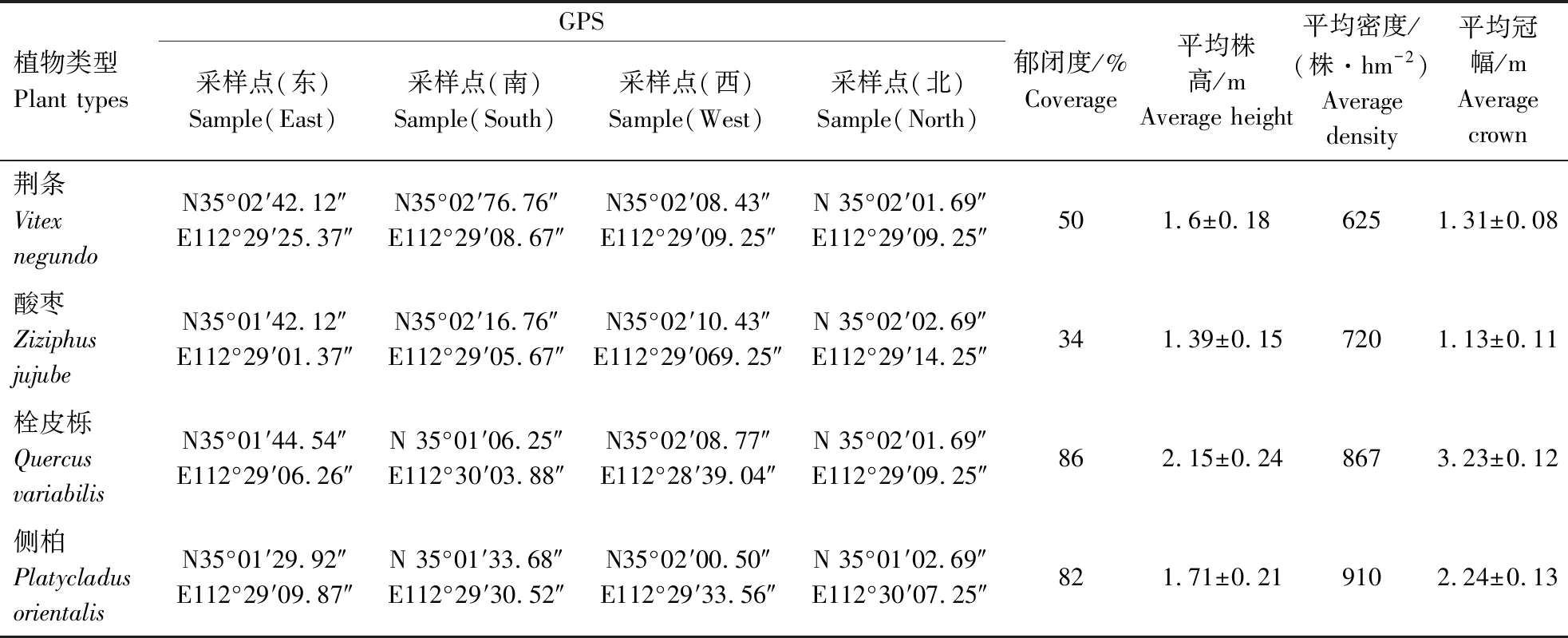
表1 样地设置基本概况Table 1 Basic overview of plot setting
1.2.2 植物叶片样品采集与测定 对于样方内4种优势植物的每一植株,均采集 5~7 片完好无病虫害的向阳成熟的叶片,将叶片置于两片湿润的滤纸之中,放入自封袋,带回实验室放入冰箱保鲜层储存[15]。测定叶干物质含量(leaf dry matter content,LDMC)、比叶面积(specific leaf area,SLA)、叶厚度(leaf thickness,LT)和光合特征值:净光合速率A、气孔导度Gs、细胞间二氧化碳浓度Ci等指标。
1.2.2.1LDMC的测定 将采集的样品叶片放入5 ℃的黑暗环境中储存12 h,取出后迅速用滤纸吸干叶片表面的水分,在1/100的电子天平上称重(饱和鲜重)。然后将叶片放入80 ℃箱内烘干24 h后取出称重(干重)。LDMC=叶片干重(mg)/叶片饱和鲜重(g)[15]。
1.2.2.2 比叶面积(SLA)SLA的测定方法用Epson Perfection V700 Photo扫描仪进行叶面积扫描,用Winfolia 2007进行叶面积的计算,得到的叶面积大小即叶大小(m2);SLA=叶片面积(m2)/叶片干重(kg)[15]。
1.2.2.3 叶厚度的测定 使用精度为0.001 mm游标卡尺,沿叶片主脉方向平均选取3个点,测量主脉一侧约0.25 cm处的厚度[16],每个叶片3个点处的厚度平均值即为叶厚度,每组测试3个叶片,其平均值即该组叶片的平均厚度。
1.2.2.4 光合特征值测定 使用CIRAS-3便携式光合测量仪,在天气晴朗的9:00—15:00之间进行叶片光合特征的测定。由于海拔差异不大,未考虑海拔影响,对4个坡向的荆条、栓皮栎、侧柏、酸枣每种植物随机选取6株,从每颗植株的同一顶端叶位向下依次选取成熟健康、无虫害的3片叶片样品测定其净光合速率A、气孔导度Gs、胞间二氧化碳浓度Ci等参数并记录,每种植物共测定18个叶片(6株×3片),并求取平均值作为最终结果。
1.2.3 土壤样品采集与测定 土壤样品采集:用无菌手套从植物根基抖落土壤,约50~100 g。将采取的土壤样品,放入无菌的自封袋密封保存,并进行编号,带回实验室进行自然风干。将土样进行风干脱水处理、研磨,分别过60目和100目筛,装入新自封袋,重新编号,以备检测使用。有机质测定采用重铬酸钾容量法-外加热法(丘林法);碱解氮采用碱解扩散法测定[16]。
1.2.4 数据处理 利用Excel和Originpro9.1对实验所得数据进行统计分析,计算和图表制作;利用SPSS22.0软件进行方差分析(ANOVA)和差异显著性检验(α差异显著性)。
2 结果与分析
2.1 优势植物不同坡向下土壤养分含量的变化
图1为4种优势植物不同坡向下根周土壤养分含量变化。结果显示,同一坡向下的4种优势植物间的土壤温度和土壤含水量无显著性差异。同一坡向下,酸枣在4种优势植物中的土壤有机质含量最高,但各植物间显著性差异不明显。而且,酸枣的土壤碱解氮含量在4个坡向下显著高于其他3种植物,尤其在北坡向上碱解氮含量高达53.95 mg·kg-1;而荆条的碱解氮含量较低,在东坡向上含量低至9.61 mg·kg-1。
2.2 优势植物不同坡向下叶面积分配性状指标变化
图2为4种优势植物不同坡向下叶面积分配性状指标变化。从图2中可以看出,同一坡向下4种优势植物间叶面积分配性状指标存在显著性差异。各坡向下,侧柏叶厚度显著高于其他3种植物,南坡厚至0.58 cm;东坡和南坡上植物间的比叶面积无显著性差异,而在西坡和北坡上侧柏的比叶面积显著高于其他3种植物,侧柏的比叶面积在北坡上高达35.48 m2·kg-1;北坡向下4种植物间的叶干物质含量无显著性差异,而在其他3个坡向下栓皮栎的叶干物质含量显著高于其他3中植物,南坡向下高达412.5 g·kg-1。
2.3 优势植物不同坡向下光合作用特征值变化
图3为4种优势植物不同坡向下光合作用特征值变化。从图3中可以看出,各个坡向下4种优势植物间光合作用特征值差异显著。酸枣和荆条的净光合速率显著高于侧柏和栓皮栎,而侧柏的净光合速率在各个坡向下都最小,特别是在南坡低至0.93 μmol·m-2·s-1;酸枣的气孔导度在各个坡向下高于其他3种植物,特别是西坡高达40.75 mmol·m-2·s-1,而侧柏则是最小。侧柏在各个坡向下的胞间CO2浓度却显著高于其他3种植物,在南坡上最大可达247.48 μmol·mol-1。
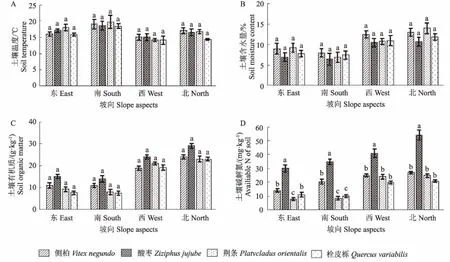
注:图中不同小写字母代表各处理平均值在P<0.05水平下差异显著。下同。Note:The different letters indicate statistical significant difference at 0.05 level.The same as below.
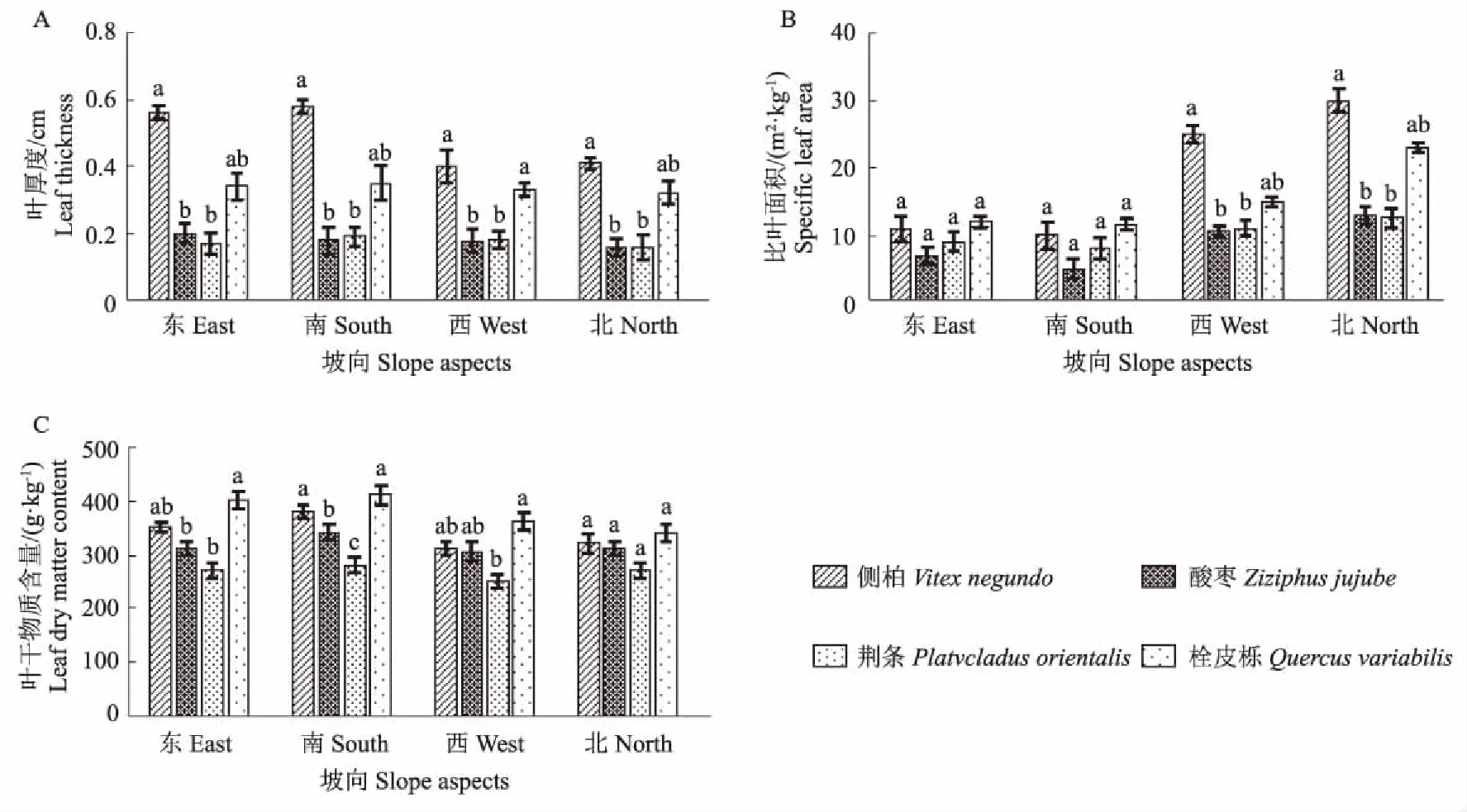
图2 4种优势植物不同坡向下叶面积分配性状变化Fig.2 Variation of leaf area distribution of four dominant plants in different slopes
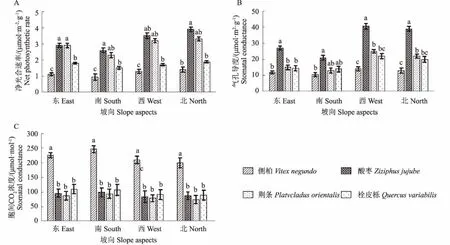
图3 4种优势植物不同坡向下光合作用特征值变化Fig.3 Changes of photosynthetic characteristics of four dominant plants in different slope aspects
3 讨论
3.1 优势植物物种间的土壤因子的差异性
植物在生长发育过程中受自身生理特性和外界环境因子的双重作用,地形因子虽然作为环境因子的一部分可以通过间接作用影响其他一些生态因子而作用于植物[5],但植物自身为进行养分吸收和生长发育需要对不同坡向具有一定的适应性以满足自身需求。研究表明,同一坡向下,酸枣在4种优势植物中的土壤有机质含量最高,但各植物间显著性差异不明显。4种优势植物不同坡向下的土壤碱解氮含量均有显著性差异,酸枣的碱解氮含量显著高于其他3种植物,尤其在北坡向上碱解氮含量高达53.95 mg·kg-1;而荆条的碱解氮含量较低,在东坡向上含量低至9.61 mg·kg-1。导致这种现象的原因可能是由于不同坡向的坡面水热条件、土壤运移和植被分布不同,致使腐殖质累积增加进而使土壤有机物发生矿化和运移,导致总有机碳的累积在不同坡向上产生差异现象[5、17-19],而且由于酸枣自身根系对土壤养分具有固定作用,从而使容易聚集土壤养分的向阴坡养分含量更高。朱云云等[20]研究发现:阳坡(东坡和南坡)在雨季时候容易发生水土流失严重和土壤矿化现象;而太行山南麓地区属于干旱半干旱地区,地势陡峭,向阳坡风化和水流侵蚀作用非常严重,土壤退化使得土壤养分本就难以富集,植被稀少,荆条在东坡向根系土壤养分含量相对较低也是由于这个原因所致[21]。
3.2 优势植物物种间的叶面积分配性状的差异性
叶面积分配功能性状作为植物的主要特性之一,主要通过叶片这个重要的植物器官进行表征[4]。研究表明,同一坡向下4种优势植物间叶面积分配性状指标存在显著性差异。各坡向下,侧柏叶厚度显著高于其他3种植物,南坡厚至0.58 cm;东坡和南坡上植物间的比叶面积无显著性差异,而在西坡和北坡上侧柏的比叶面积显著高于其他3种植物,侧柏的比叶面积在北坡上高达35.48 m2·kg-1;植物的叶干物质含量除了在北坡向无显著性差异外,其他3个坡向均存在显著性差异,在东、南、西坡向上栓皮栎的叶干物质含量显著高于其他3种植物,最大为412.5 g·kg-1。叶干物质含量与植物抵御外界的能力呈正相关关系,是植物适应外界环境变化的关键性状[22-23]。不同植物物种间的叶干物质含量发生变化可能是由于不同物种受到不同环境条件胁迫下所表现出的对生物量分配策略的不同[5],本研究中栓皮栎的叶干物质含量显著高于其他植物物种,这与胡耀升等[24]的植物内部的水分会随着叶干物质含量的增加而减少损失的研究相一致。南坡光照充足,但由于土壤含水量较少,叶片出于保护增加了细胞密度,叶片较厚;而北坡光照资源欠缺,但水资源充足,细胞体积和数目增加,促进了叶面积的扩展,叶片较薄,所以侧柏的叶厚度在南坡最大,而生长于北坡的侧柏比叶面积较大。植物的比叶面积反映植物获取和利用资源的能力,与植物的生存对策有密切的联系。比叶面积高,代表其叶片的光捕获面积高,而比叶面积较低的植物,叶的大部分物质用于构建保卫构造, 常形成厚度较大而面积较小的叶片[25-26]。
3.3 优势植物物种间的光合作用特征的差异性
光合作用特征代表的是植物进行光合作用积累有机物的指标[27-28]。本研究表明,各个坡向下4种优势植物间光合作用特征值差异显著。酸枣和荆条的净光合作用速率显著高于侧柏和栓皮栎,侧柏的净光合速率在各个坡向下都最小,特别是在南坡低至0.93 μmol·m-2·s-1;酸枣的气孔导度在各个坡向下高于其他3种植物,特别是西坡高达40.75 μmol·m-2·s-1,而侧柏则是最小。侧柏在各个坡向下的胞间CO2浓度却显著高于其他3种植物,在南坡上最大可达247.48 μmol·mol-1。产生这种现象的原因是由于北坡的光照强度不足,植物呼吸作用弱,叶片气孔扩散阻力减少,叶片外面的空气和叶绿体内的羧化部位之间的浓度梯度越大,导致从叶外空气到羧化部位之间的扩散阻力越小,进入细胞的CO2越多,叶片净光合速率和气孔导度越高[29-30]。本研究中,东坡和南坡尽管光照充足会增加植物的总光合作用速率,但是蒸腾作用和呼吸作用消耗相应增加,叶片出于保护自身会降低气孔导度和净光合速率,一般形成的叶片较厚,所以适合南坡的净光合速率最小的侧柏等乔木植物的生长;而北坡光照资源欠缺,植物为保持自身所需能量平衡,必须降低呼吸作用,从而降低叶片气孔阻力,增加气孔导度和净光合作用速率,一般植物叶片较大而薄,适合酸枣等灌木类生长。
4 结论
对太行山南麓4种优势植物间不同坡向下的叶功能性状与土壤养分含量进行了研究。结果表明:1)植物自身对外界环境具有强大的适应性,会根据自身需求采取最有利的生长条件;2)酸枣等灌木类植物在光照资源欠缺的北坡更易聚集养分;3)侧柏为适应环境变化满足自身生长叶片在南坡较厚,而在北坡比叶面积却较大;4)植物的光合作用特征指标表明:侧柏为适应光照充足的南坡保护自身而降低气孔导读和净光合速率,而胞间CO2浓度变化是相反的。
猜你喜欢
花卉(2024年1期)2024-01-16 11:33:50
林业科学研究(2023年4期)2023-04-29 00:44:03
新农民(2022年22期)2022-11-07 23:38:06
今日农业(2022年13期)2022-09-15 01:18:00
中成药(2021年5期)2021-07-21 08:38:46
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
星星·诗歌原创(2021年3期)2021-06-01 09:13:23
新疆农业科学(2020年1期)2020-02-14 03:22:46
河北果树(2020年1期)2020-02-09 12:31:42
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
