
我国草地贪夜蛾的生物学、生态学和防治研究概况与展望
2020-07-08 00:53谷少华高希武
昆虫学报 2020年5期
梁 沛, 谷少华, 张 雷, 高希武
(中国农业大学昆虫学系, 北京 100193)
草地贪夜蛾Spodopterafrugiperda原产美洲,由于其具有寄主范围广、食量大、繁殖和迁飞能力强等特点,已成为世界性重大害虫。自2018年12月11日从缅甸侵入我国云南以来(Sunetal., 2019),给我国的粮食安全造成严重威胁。
知己知彼,百战不殆。对草地贪夜蛾深入系统的研究是实现其可持续治理的重要前提。由于草地贪夜蛾在国外发生历史较长,国际上对该害虫已经进行了广泛而深入的研究。以“Spodopterafrugiperda”为主题词在Web of Science数据库中查询,在1970-2019年共有12 362篇文献,1990年以来平均每年发文量在250篇左右。研究内容涉及昆虫学、细胞生物学、生化及分子生物学、遗传学、植物学、病毒学等80多个学科。
草地贪夜蛾入侵我国之前,国内相关研究甚少。2005-2018年共发表16篇相关论文,年均仅1.14篇,且多是以草地贪夜蛾细胞系为材料,研究动物毒素、植物次生代谢产物或杀虫剂对细胞的毒性(王文祥等, 2011; 汪丽等, 2013; 杨韵等, 2017),以草地贪夜蛾种群为对象的研究论文基本没有。自2018年底草地贪夜蛾入侵我国之后,我国迅速加大了对该害虫的研究,有关草地贪夜蛾的论文呈井喷式发表。截止2020年1月底,以“草地贪夜蛾”为关键词,在中国知网(CNKI)中共检索到研究性论文和文献综述242篇,仅2019年就发表223篇中文论文和10篇左右的英文论文。其中,中国农业科学院植物保护研究所发表了近60篇,占草地贪夜蛾论文总数的71.6%,远超分别位列第二和第三的全国农业技术推广服务中心和华南农业大学。这些论文主要发表在国内10种期刊上,其中《植物保护》、《环境昆虫学报》、《中国生物防治学报》、《西南大学学报(自然科学版)》和《中国植保导刊》5种期刊的发文数量占总数的65%。
从已发表的论文来看,目前国内的研究主要集中在草地贪夜蛾的生物学、生态学、生物防治、化学防治、基因组学及综合防控等方面。本文拟对目前我国草地贪夜蛾的研究现状做一梳理,以期为国内同行进一步深入系统地研究这一重大入侵害虫、实现其可持续防控提供参考。
1 分类鉴别和生物学研究
这方面研究主要涉及分类鉴定、生物型鉴定、为害作物调查及饲养条件探索等。
1.1 分类鉴定
作为一种全新的外来入侵害虫,从科研人员、农业技术推广人员到普通农户,对草地贪夜蛾都缺乏了解,绝大部分人根本不认识。而害虫种类的正确鉴定和识别是对其进行有效防治的重要前提。张智等(2020)通过对灯诱监测过程中诱集到的不易区分的草地贪夜蛾、旋歧夜蛾Anartatrifolii、陌夜蛾Tracheaatriplicis、甘蓝夜蛾Mamestrabrassicae和斜纹夜蛾Spodopteralitura5种夜蛾成虫的形态分析,认为其翅、前足基部及雄性外生殖器的形态可作为准确识别这5种夜蛾成虫的关键分类特征。李国平等(2019b)对黄淮海夏玉米区混合发生的草地贪夜蛾、甜菜夜蛾Spodopteraexigua、棉铃虫Helicoverpaarmigera、玉米螟Ostrinianubilalis和劳氏黏虫Leucanialoreyi5种幼虫的形态做了详细比较。陈琦等(2020)则系统比较了在玉米灌浆期常与草地贪夜蛾混合发生的黏虫Mythimnaseparata、劳氏黏虫和棉铃虫等害虫的卵、幼虫、蛹和成虫的外部形态特征,为玉米灌浆期草地贪夜蛾的准确鉴别和田间调查提供了依据。另外,董前进等(2019)提出了一种快速鉴别草地贪夜蛾蛹及成虫性别的简易方法;张智等(2019b)描述了草地贪夜蛾覆毛卵块和绒茧蜂Apantelesspp.茧块的识别特征。这些都为从外部形态上准确鉴别混合发生的形态近似的害虫提供了重要借鉴。Jing等(2019)则利用PCR技术,建立了基于细胞色素氧化酶 Ⅰ(cytochrome oxidase subunit Ⅰ, COⅠ)和磷酸丙糖异构酶(triosephosphate isomerase, Tpi)基因的鉴别草地贪夜蛾、甜菜夜蛾、黏虫和斜纹夜蛾的方法。上述基于形态学和基因序列差异的鉴定方法,为草地贪夜蛾的识别提供了重要技术支撑,但其效率相对比较低。于业达等(2019)基于不同地域、不同发育阶段的草地贪夜蛾及相似物种的图片数据库,利用三通道T型深度卷积神经网络(tubelets with convolutional neural network, T-CNN)对数据进行分类,建立了一种基于深度学习的草地贪夜蛾智能识别技术,平均识别率达到97%,使草地贪夜蛾的快速识别和种群动态自动监测成为可能。如果能进一步完善并尽快应用到田间草地贪夜蛾的自动监测,将会极大降低人工监测的工作强度。
1.2 生物型鉴定
草地贪夜蛾有玉米型(corn strain)和水稻型(rice strain)两种生物型(Pashley and Martin, 1987)。不同生物型其生物学特性、为害特性以及对杀虫剂的抗性等均存在显著差异。因此,明确入侵我国的草地贪夜蛾的生物型至关重要。张磊等(2019)利用COⅠ和Tpi两个基因片段对采自我国13个省(市、自治区)131个县(市、区)的318份草地贪夜蛾样品进行检测,认为入侵我国的草地贪夜蛾很可能是一种特殊的玉米型,即是由水稻型母本和玉米型父本的杂交后代长期演化形成的,且玉米型核基因组占据主导地位。但利用同样的两个分子标记,不同研究者对不同地区草地贪夜蛾生物型的鉴定结果存在较大差异。如陈冬平等(2020)利用COI-A,COI-B和COI-C标记鉴定认为入侵广州的草地贪夜蛾为水稻型,甚至鉴定到了草地贪夜蛾与黏虫的杂合子;而利用Tpi基因则鉴定为玉米型。王佳丽等(2020)同样利用COⅠ和Tpi基因对采集自广东、广西和湖南3个省(区)的样本分析认为,除湖南张家界种群中2个样品为COⅠ玉米型外,其他种群均为COⅠ水稻型。唐运林等(2019)对入侵重庆不同地区的草地贪夜蛾利用COⅠ基因鉴定认为全为水稻型,而利用Tpi基因鉴定既有水稻型又有玉米型。徐丽娜等(2019b)基于COⅠ的分析结果显示安徽省草地贪夜蛾样本中86.23%是水稻型,13.77%是玉米型,而基于Tpi基因序列分析认为所有样品均为玉米型。Zhang等(2019)对采自我国16个省的105个草地贪夜蛾样本进行基因组重测序分析发现,所有样本的遗传背景均由70%以上的玉米型、不到15%的水稻型和15%的杂合型组成,并进一步验证发现利用Tpi基因的鉴定结果与基因组重测序的结果高度吻合,可用于草地贪夜蛾两种生物型的鉴定;而基于COⅠ基因的鉴定结合与基因组重测序鉴定结果无相关性,因而不适合用于生物型鉴定。可见,分子标记的选择对生物型鉴定的结果影响极大。建议今后统一采用Tpi基因对草地贪夜蛾的生物型进行鉴定。
1.3 为害作物调查
据文献报道,草地贪夜蛾可为害76科353种植物(Montezanoetal., 2018),尤其可对玉米、水稻、高粱、花生、棉花、大豆和甘蔗等粮食作物造成严重为害。草地贪夜蛾入侵我国后,虽然主要为害玉米,但在其他作物上的发生情况也一直受到密切关注。2019年4月,太红坤等(2019)发现草地贪夜蛾在云南德宏州为害甘蔗苗。随后,不断有草地贪夜蛾为害甘蓝(刘银泉等, 2019)、马铃薯(赵猛等, 2019)、花生(何莉梅等,2020)、小麦(徐丽娜等, 2019a; 李艳朋等, 2020)、冬粉薯(周上朝等, 2020)、青稞、燕麦和糜子(赵雪晴等, 2020)等多种作物的报道。另外,还有为害茶树和烟草的可能(徐蓬军等, 2019; 孙晓玲等, 2020)。据姜玉英等(2019)报道,草地贪夜蛾在我国为害的作物已达15种,另外还可为害皇竹草Pennisetumsinese、马唐Digitariasanguinalis、牛筋草Eleusineindica和苏丹草Sorghumsudanense等禾本科杂草。随着该害虫在我国的定殖,其为害的作物范围还可能进一步扩大。因此,在加强玉米田草地贪夜蛾防控的同时,也要严密监测其在其他粮食和经济作物上的发生和为害情况,严防对其他作物的安全生产造成威胁。
1.4 温度适应性研究
昆虫对低温的适应能力决定了昆虫的生长发育和地理分布,进而影响其为害范围和为害程度。何莉梅等(2019)测定了草地贪夜蛾各发育阶段的发育起点温度、有效积温和生命表参数,并构建了发育历期的预测模型,研究结果为预测草地贪夜蛾的发育历期、发生区域、发生期和发生世代提供了重要依据。张智等(2019b)测定发现草地贪夜蛾不同虫态的过冷却点和体液冰点存在显著差异,其中卵的过冷却点和体液冰点最低,分别为-25.45℃和-24.70℃,而6龄幼虫的过冷却点和体液冰点最高,分别为-7.35℃和-2.07℃;幼虫的过冷却点和体液冰点均随龄期增加而升高;雌雄成虫的过冷却点无显著差异,但雌成虫体液冰点显著低于雄成虫。总体来看,草地贪夜蛾具有较强的抗寒能力。另外,张悦等(2020)和谢殿杰等(2020)分别研究了取食不同食物和不同饲养温度对草地贪夜蛾过冷却点和体液冰点的影响。上述研究为划分草地贪夜蛾的越冬区和监测预警提供了科学依据。
1.5 其他生物学特性
房敏等(2020)比较了成虫期补充不同营养对草地贪夜蛾繁殖力的影响,发现补充10%蜂蜜水或10%蔗糖水的雌成虫的产卵期及寿命均显著长于其他处理组,尤其是补充10%蜂蜜水的产卵量及卵孵化率均最高,分别为983.2粒和98.07%。金涛等(2020)在室内条件下研究了草地贪夜蛾雌成虫的产卵节律,发现平均单雌总产卵量在1 000粒左右,其中羽化后7 d内的产卵量占总产卵量的68.1%,最高日产卵量可达229.3粒/头;产卵主要在晚上20∶00-次日5∶00时进行;与心叶相比,草地贪夜蛾更偏好在玉米老叶上产卵。 王道通等(2020)研究了草地贪夜蛾不同龄期的幼虫在不同饲养密度下的自相残杀行为,认为随着龄期和密度增大,其自相残杀行为加剧。另外,王世英等(2019)和苏湘宁等(2019)研究了草地贪夜蛾人工饲料和饲养技术。这些都为草地贪夜蛾的室内大量饲养提供了有益参考。
2 生态学研究
这方面的研究主要集中在草地贪夜蛾的扩散为害规律、预测预报技术、潜在地理分布及迁飞规律等方面。
2.1 扩散为害规律研究
姜玉英等(2019)通过全国范围的系统监测,全面总结了2019年草地贪夜蛾在我国的侵入及扩散为害规律,结果显示草地贪夜蛾主要为害玉米;在长江以南地区普遍发生,并由南至北逐步扩散,虫口密度由南至北逐步递减;发生时间西部早于东部,虫口密度西部大于东部;并根据发育起点温度和有效积温推算出草地贪夜蛾在我国从南到北最多每年可发生6~8代,最少2~3代;在此基础上提出未来应加强对缅甸、泰国和越南等东南亚国家虫源的监测,在南方种植春玉米以诱集草地贪夜蛾,延缓其北迁时间、减轻北迁虫量的策略。李向永等(2019)和卢辉等(2019)分别调查了缅甸北部和越南北部草地贪夜蛾的发生情况,为我国草地贪夜蛾的监测预警和防控提供了参考。
2.2 测报技术研究
测报技术的研究是准确预测预报害虫发生动态的重要基础。目前对草地贪夜蛾的测报技术从不同层次做了研究。赵胜园等(2019a)研究发现其卵巢发育分级与卵巢管长度和宽度符合3次多项式方程关系,而与卵粒直径及抱卵量符合2次方程关系,在此基础上提出了草地贪夜蛾卵巢发育分级与繁殖潜力的预测方法。何沐阳等(2019)建立了玉米苗期有虫株率与幼虫密度的指数函数模型,并提出了以这两个参数为指标的草地贪夜蛾发生程度分级标准。刘杰等(2019)提出了针对草地贪夜蛾卵、幼虫、蛹和成虫等不同虫态的种群监测方法,系统介绍了田间调查技术。另外,杨现明等(2020a, 2020b)研究了小麦田和大麦田草地贪夜蛾的空间分布和抽样技术,孙小旭等(2019)研究了玉米田草地贪夜蛾的空间分布型和抽样技术,和伟等(2019)报道了草地贪夜蛾种群的性诱测报方法。上述研究为系统建立草地贪夜蛾的虫情调查、种群发生期和发生量等测报技术体系提供了重要支撑。但基于这些技术的预测预报均要通过人工田间调查来实现,技术要求高,又费时费力,且只在小范围适用。如何实现基于自动化、智能化的大区域、大尺度下草地贪夜蛾种群动态的准确监测和预报,还有待进一步加强研究。
2.3 潜在地理分布研究
草地贪夜蛾作为一种新的外来入侵害虫,尽快研究明确其在我国可能的地理分布范围,对于宏观尺度下草地贪夜蛾的监测预警和可持续防控策略的制订都具有重要意义。秦誉嘉等(2019)、林伟等(2019)和Wang等(2020)分别利用MaxEnt模型预测了草地贪夜蛾在我国的潜在地理分布,并且均划分出了高度适生区、中度适生区、低度适生区和非适生区,明确了草地贪夜蛾在我国绝大部分区域均可分布。其中,秦誉嘉等(2019)认为除黑龙江和吉林外,其他省份均可分布;而林伟等(2019)认为在全国各省区均可分布。另外,这3项研究在适生区的划分上也有一定差异,有待进一步研究明确。总体来看,这3项研究都只是初步研究,在预测时只考虑了气候因素,如温度、降雨量等。而草地贪夜蛾以玉米为主要寄主,如果在预测时能进一步考虑土地利用率、作物种类及种植模式等影响因素,预测结果可能会更准确。
2.4 迁飞路线研究
草地贪夜蛾具有很强的迁飞能力,这是其迅速扩散、造成大面积为害的重要原因。明确其进入我国的路径及在国内的主要迁飞路线,对于该害虫的监测预警和有效防控同样至关重要。中国农业科学院植物保护研究所吴孔明院士团队和南京农业大学胡高教授团队联合对此进行了深入研究,结果表明草地贪夜蛾可通过东、西两条路径进入我国并迁飞至北方玉米主产区:西线路径是从缅甸进入云南,经贵州进入四川、重庆、河南以至陕西、山西;东线路径则是从越南、老挝、泰国进入广东和广西,依靠强劲的西南风,直达长江流域和淮河流域,进入我国玉米主产区华北平原,甚至东北平原(吴秋琳等, 2019; Wuetal., 2019; Lietal., 2020)。这为合理安排监测点,严密监控草地贪夜蛾的迁飞情况提供了重要依据。
3 生物防治研究
有关草地贪夜蛾的天敌资源及其在生物防治中的应用,已经有多篇综述对国内外的文献做了较为全面的总结(陈万斌等, 2019; 唐璞等, 2019; 颜珣等, 2019; 张维等, 2019)。
国内目前有关草地贪夜蛾生物防治方面的研究主要集中在对草地贪夜蛾有控制作用的本地天敌种类调查及生防功能评价两个方面。随着草地贪夜蛾在国内的快速扩散,各地迅速展开了对本地天敌资源的调查。目前共发现捕食性天敌10种,其中鞘翅目和半翅目各4种,脉翅目和铗尾目各1种;寄生性天敌共12种,其中膜翅目11种,双翅目1种;昆虫核型多角体病毒5种,昆虫病原真菌4种(表1)。
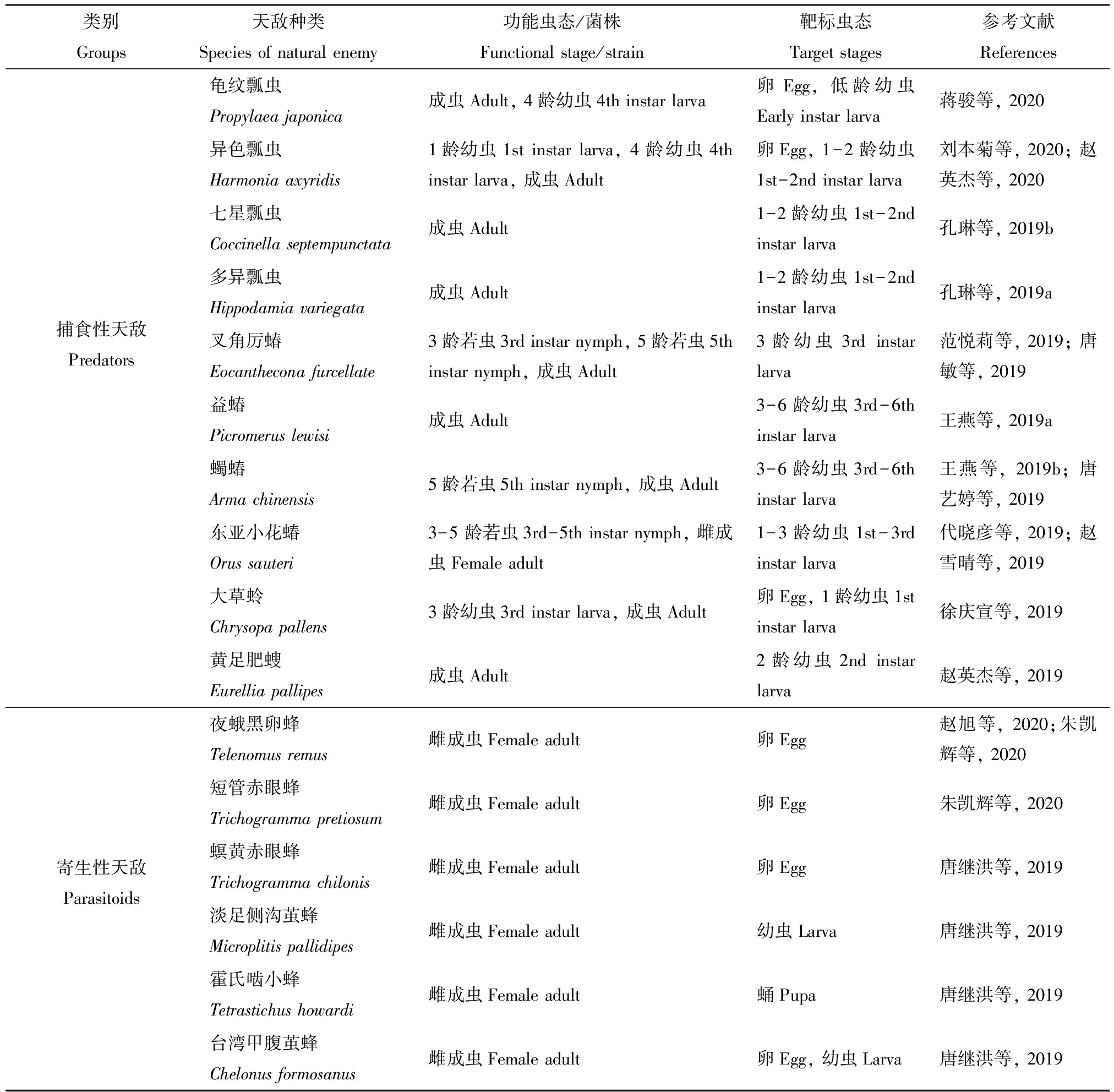
表1 已报道的对草地贪夜蛾有控制作用的本土天敌Table 1 Reported local natural enemies used for control of the fall armyworm, Spodopterar frugiperda in China
续表1 Table 1 continued

类别Groups天敌种类Species of natural enemy功能虫态/菌株Functional stage/strain靶标虫态Target stages参考文献References螟甲腹茧蜂Chelonus munakatae雌成虫Female adult卵Egg李芬等, 2019菜粉蝶盘绒茧蜂Cotesia glomerata 雌成虫Female adult卵Egg宁素芳等, 2019半闭弯尾姬蜂Diadegma semiclausum雌成虫Female adult卵Egg宁素芳等, 2019斯氏侧沟茧蜂Microplitis similis雌成虫Female adult2-4龄幼虫2nd-4th instar larva陈壮美等, 2019斑痣悬茧蜂Meteorus pulchricornis雌成虫Female adult幼虫Larva尚丹等, 2019日本追寄蝇Exorista japonica雌成虫Female adult幼虫Larva宁素芳等, 2019核型多角体病毒棉铃虫NPVHelicoverpa armigera NPV-幼虫Larva张海波等, 2020Nucleopolyhedrovirus甜菜夜蛾NPVSpodoptera exigua NPV-幼虫Larva(NPV)甜菜夜蛾NPVSpodoptera exigua NPV-幼虫Larva甘蓝夜蛾NPVMamestra brassicae NPV-幼虫Larva亚洲玉米螟NPVOstrinia furnacalis NPV-幼虫Larva昆虫病原真菌Entomopathogenic fungus玫烟色虫草Cordyceps fumosorosea SCAU-IFCF01菌株SCAU-IFCF01 strain幼虫Larva雷妍圆等, 2020金龟子绿僵菌Metarhizium anisoplaeCQMa421菌株CQMa421 strain低龄幼虫Early instar larva, 蛹Pupa, 卵Egg彭国雄等, 2019球孢白僵菌Beauveria bassianaZJU435菌株ZJU435 strain低龄幼虫Early instar larva, 蛹Pupa, 卵Egg彭国雄等, 2019莱氏绿僵菌Metarhizium rileyi ZYSP190701菌株ZYSP190701 strain幼虫Larva郑亚强等, 2019
针对上述发现的本土天敌,进一步研究了在室内不同条件下对不同发育阶段草地贪夜蛾的控制潜能,如捕食性天敌的捕食功能反应、寄生性天敌的寄生率、昆虫病毒和病原真菌的致毒力等,但真正对田间防治效果的研究相对较少。因此,进一步通过田间试验,评价、筛选出在不同地区、不同气候条件下对草地贪夜蛾具有较好防控作用的天敌种类,并加快其产业化是今后一段时期研究的重点。
在生物源农药对草地贪夜蛾的防治效果方面也做了一些研究,如林素坤等(2020)在测定印楝素对草地贪夜蛾室内毒力的基础上,进一步评价了其在田间的防治效果;赵胜园等(2019b)测定了10种常用生物源和生物杀虫剂对草地贪夜蛾的室内毒力,认为多杀霉素、阿维菌素、苏云金芽孢杆菌Bacillusthuringiensis(Bt)和球孢白僵菌Beauveriabassiana有较好的毒力。这为将生物源和生物杀虫剂用于防治草地贪夜蛾奠定了一定基础,但还有待进一步通过田间试验进行验证。此外,国外实践证明,种植转Bt基因抗虫玉米是有效防控草地贪夜蛾的一项重要措施。李国平等(2019a)室内测定发现,Cry1Ab, Cry1Ac, Cry1F, Cry2Ab及Vip3A 5种Bt蛋白对草地贪夜蛾幼虫均有较高的毒力。张丹丹和吴孔明(2019)进一步测定了两种国产转基因玉米Bt-Cry1Ab和Bt-(Cry1Ab+Vip3Aa)对草地贪夜蛾的活性,发现两种玉米均能高效表达目标杀虫蛋白,并且对草地贪夜蛾具有很强的杀虫活性。这为国内通过推广种植转Bt抗虫玉米防控草地贪夜蛾提供了重要的实验依据。
4 化学防治研究
化学防治是对暴发性害虫进行应急防控的最重要也是最有效的措施。由于草地贪夜蛾刚入侵我国,因此国内并没有登记用于防治该虫的杀虫剂。针对这种情况,农业农村部于2019年6月3日发布了关于做好草地贪夜蛾应急防治用药有关工作的通知(http:∥www.gov.cn/xinwen/2019-06/10/content_5398774.htm),在专家组充分论证的基础上,推荐了25种用于草地贪夜蛾应急防控的杀虫药剂,其中19种为化学杀虫剂。与此同时,国内相关研究单位也对现有可用于鳞翅目害虫防治的杀虫药剂进行了大量筛选。到目前为止,国内对于草地贪夜蛾的化学防治研究主要集中在对草地贪夜蛾高活性杀虫药剂的室内筛选,包括杀幼虫的药剂(鲁艳辉等, 2019; 尹艳琼等, 2019)和具有杀卵活性的药剂的筛选(林玉英等, 2019; 王芹芹等, 2019a),以及对草地贪夜蛾田间药效评价(金涛等, 2019; 闫文娟等, 2019; 赵胜园等, 2019c; 胡飞等, 2020)两个方面。不同地区、不同研究机构仅2019年就发表此类论文近30篇,对指导对当地合理用药、有效防治草地贪夜蛾起到了积极作用。但不足之处是重复研究较多,很多对鳞翅目害虫防效较好的药剂如甲维盐、茚虫威、氯虫苯甲酰胺等在多个研究中重复出现,造成了一定程度的资源浪费。
除了药剂筛选,在药剂高效使用技术方面也已经做了一些工作。如朱峰等(2019)研究发现使用低容量连杆多喷头喷雾对草地贪夜蛾有良好的防治效果;王磊等(2019)评价了植保无人机减量施药情况下对草地贪夜蛾的防治效果;刘妤玲等(2019)研究了超低容量液剂对玉米田草地贪夜蛾的防治效果;杨帅等(2020)发现同样是四氯虫酰胺,在防治效果相当的情况下,喇叭口点施要比整株喷雾减药50%。但总体来看,针对草地贪夜蛾的高效、精准、减量施药技术还比较欠缺。最近农业农村部印发了《2020年全国草地贪夜蛾防控预案》(http:∥www.moa.gov.cn/xw/bmdt/202002/t20200221_6337551.htm),继续推荐了28种应急防控药剂,其中22种为化学杀虫剂,包括8种单剂和14种混剂,这些都是目前对鳞翅目害虫防效优异的药剂。但如何才能真正发挥其在草地贪夜蛾防治中的作用,则需要尽快研发与草地贪夜蛾发生为害特点相适应的高效对靶施药技术。
在草地贪夜蛾的化学防治中,一个不可忽视的问题就是杀虫药剂的抗性问题。虽然草地贪夜蛾才刚刚进入我国,我们也才开始对其进行化学防治,但在进入我国之前由于长期的化学防治,导致其已经对有机磷类、氨基甲酸酯类及拟除虫菊酯类等多种传统杀虫药剂产生了高水平抗性,对氯虫苯甲酰胺等新型双酰胺类杀虫剂也产生了抗性。有关草地贪夜蛾对化学杀虫剂的抗性现状已经有多篇综述(李永平等, 2019; 王芹芹等, 2019b; 吴超等, 2019; 吴益东等, 2019),这里不再赘述。在入侵我国多地的草地贪夜蛾种群中,均已检测到高频率的与有机磷类、氨基甲酸酯类和拟除虫菊酯类杀虫剂抗性相关的靶标基因突变。如Zhang等(2019)通过对采自我国16个省的105个个体的基因组重测序发现,在有机磷和氨基甲酸酯类的作用靶标乙酰胆碱酯酶上存在两个已经报道的与抗性相关的突变A201S和F290V,其突变频率分别高达17.1%和58.2%;同时还检测到与菊酯类杀虫剂抗性相关的神经膜钠离子通道的L932F突变,其频率高达45.5%。我们也检测到与双酰胺类杀虫剂抗性相关的鱼尼丁受体的I4790K突变(未发表数据),虽然频率很低,但也应该引起高度重视。另外,Liu等(2019)通过对草地贪夜蛾基因组测序发现,其细胞色素P450和谷胱甘肽S-转移酶两个与杀虫剂代谢抗性密切相关的基因家族出现大量扩张。这些均说明进入我国的草地贪夜蛾本身已经携带有高频率的抗性基因,加之其高繁殖率等生物学特性以及目前我国以化学防治为主的控制策略,如果不尽快制订并实施科学合理的综合治理和抗性治理策略,草地贪夜蛾的抗药性问题将很快凸显,严重影响化学防治效果。
5 性信息素及性引诱剂研究
常亚军等(2019)对有关草地贪夜蛾性信息素的组成及其对草地贪夜蛾引诱效果的研究做了简要综述,发现不同地区草地贪夜蛾的性信息素组分及比例存在差异。因而要开发普适高效的性信息素引诱剂有一定难度,需要进行更多细致、系统的研究。江南纪和王琛柱(2019)在对草地贪夜蛾性信息素通讯研究现状综述的基础上提出了3点建议:(1)尽快鉴定入侵我国草地贪夜蛾种群的雌性性信息素并用于其种群动态监测及防治;(2)研究雄蛾对性信息素的嗅觉编码机制;(3)研究玉米型和水稻型草地贪夜蛾的行为隔离程度及其演化结局,以更好地服务于草地贪夜蛾的防控实践。
车晋英等(2020)发现4家不同公司的性诱剂产品在田间的引诱效果存在明显差异;沈嘉彬等(2019)测试了6种不同配方的性信息素诱芯对草地贪夜蛾的田间诱捕效果,发现有效成分顺-9-十四乙酸酯∶顺-7-十二乙酸酯=96.6∶3.4(m/m)、含量为3.0 mg的诱芯引诱效果较好。杨留鹏等(2019)则从9种诱芯和5种类型的诱捕器中筛选出了对草地贪夜蛾雄蛾诱捕效果最佳的诱芯和诱捕器组合,为草地贪夜蛾的综合防控提供了更多选择。
在草地贪夜蛾的化学感受机制方面也做了一些工作。葛文超等(2019)描述了草地贪夜蛾幼虫头部感受器的种类、分布和数量特征;刘莹等(2019)基于草地贪夜蛾的基因组测序结果,对其化学感受相关基因家族的进化进行了分析;黄钧鸿等(2019)则对草地贪夜蛾和斜纹夜蛾嗅觉、味觉相关的基因进行了比较分析。这些都为进一步阐明草地贪夜蛾的化学通讯机制,进一步研发更加高效、专一的草地贪夜蛾的性诱剂、食诱剂等提供了一定基础。但这些都只是一些初步研究,尚有待更为深入系统地研究,以揭示草地贪夜蛾化学感受的分子机制,为应用于生产实践提供理论基础。
6 综合防控技术研究
虽然目前对草地贪夜蛾的防治策略是以化学防治为主的应急防控,但随着时间推移,该害虫必将成为我国的一个常发性害虫,因此以预防为主的综合防控将成为主要策略。在这方面,徐汉虹等(2020)对昆虫种群遗传调控技术(如性连锁平衡致死系统、构建杂交不育平衡致死品系)和干扰昆虫生殖的相关技术(如控制昆虫性别、干扰昆虫生长发育的关键基因、调整性信息素比例等)进行了综述,并展望了这些技术在草地贪夜蛾防控中的应用前景;Ren等(2019)则提出了以表达多个作用靶标不同的Bt杀虫蛋白基因为关键,以沉默生长发育、解毒代谢、消化和肠道防御等关键基因为核心的多基因聚合和沉默(multiple genes pyramiding and silencing, MGPS)策略,为构建同时表达多个Bt杀虫蛋白和dsRNA的转基因玉米,克服对单价转基因抗虫玉米的抗性,实现对草地贪夜蛾的可持续控制提供了思路。
吴孔明(2020)则从更加宏观的尺度提出了我国草地贪夜蛾防控工作分“两步走”的策略:第一步是在近期实施传统的综合防治技术体系以解决短期内草地贪夜蛾的应急管控问题;第二步是通过现代农业信息技术和生物技术的创新与应用,力争在3~5年内构建和实施以精准监测预警、迁飞高效阻截和种植Bt玉米为核心的综合防治技术体系,实现对草地贪夜蛾的低成本和绿色可持续控制。
7 小结与展望
总体来看,在草地贪夜蛾进入我国的短短一年内,我国的植保科技人员在前期几乎无任何基础的情况下,迅速开展工作,在草地贪夜蛾的生物学、生态学、生物防治、化学防治及性引诱剂等方面做了大量工作,取得了较大进展,为进一步开展更为深入、系统的研究工作奠定了坚实基础。但其中也存在一些问题,如有些工作做得比较仓促,不够细致,缺乏深度;有些工作则是简单重复,缺乏创新,等等。希望今后的研究更加认真细致、深入系统,真正为我国草地贪夜蛾的可持续防控贡献力量。
草地贪夜蛾作为全球预警的重大迁飞性害虫,已在我国南方定殖,同时境外虫源也将持续迁入,防控形势更加严峻。因此,今后一段时间建议加强以下几个方面的工作,为实现草地贪夜蛾的可持续防控奠定坚实基础:
(1)在加强玉米田草地贪夜蛾防控的同时,继续严密监控草地贪夜蛾对其他作物的为害情况,严防其对其他作物造成严重为害;
(2)进一步完善昆虫智能识别技术并研发相关产品,尽快实现草地贪夜蛾种群动态的自动监测和预报,为准确及时的防控决策提供依据;
(3)针对已发现的本地天敌,筛选确定具有较好应用前景的种类,尽快实现产业化,同时加快转Bt抗虫玉米的商品化,以减少化学杀虫剂的使用强度;
(4)将草地贪夜蛾纳入已有害虫抗药性监测网络,密切监测其对主推杀虫剂的抗性水平及抗性基因频率,并根据监测结果,因地制宜,及时调整化学防控策略,避免抗药性加剧;
(5)尽快研发高效性诱剂和食诱剂产品并用于草地贪夜蛾的种群动态监测和综合防控;
(6)加强生物学基础理论研究,以基因组数据为基础,应用多组学的理论和技术,研究明确调控草地贪夜蛾生长、发育、生殖、化学感受等重要生命过程的分子机制及关键基因,为发展基于昆虫种群遗传调控的防治新技术奠定理论基础;
(7)加强草地贪夜蛾对主要化学杀虫剂和不同类型Bt杀虫蛋白产生抗性的分子机制研究,为延缓其对杀虫剂和转基因抗虫玉米的抗性,实现草地贪夜蛾的绿色可持续控制提供理论指导。
猜你喜欢
少儿科学周刊·儿童版(2021年21期)2021-12-11
云南农业(2021年11期)2021-11-12
今日农业(2020年13期)2020-12-15
幼儿100(2020年31期)2020-11-18
小天使·二年级语数英综合(2019年10期)2019-11-08
疯狂英语·初中版(2019年4期)2019-09-10
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
小太阳画报(2018年6期)2018-05-14
