
我国草地贪夜蛾田间种群有机磷和氨基甲酸酯类杀虫剂靶标基因ace-1的基因型和突变频率
2020-07-08 00:53龚丽凤王欢欢谷少华高希武
昆虫学报 2020年5期
李 妍, 龚丽凤, 王欢欢, 李 曦, 孙 歌,谷少华,*, 梁 沛, 高希武
(1. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京100193; 2. 中国农业大学昆虫学系, 北京 100193)
草地贪夜蛾Spodopterafrugiperda,又称秋粘虫,隶属于鳞翅目(Lepidoptera)夜蛾科(Noctuidae)灰翅夜蛾属Spodoptera,原产于美洲热带和亚热带地区,最早记载于1797年,1856年报道为害棉花,1917年报道了其防治方法(Murúaetal., 2009; 李永平等, 2019)。草地贪夜蛾是一种杂食性重大农业害虫,在美洲记载的寄主植物多达76科353种,能够对玉米、小麦、水稻、高粱、甘蔗、棉花等作物造成重大产量和经济损失,如可造成美国玉米产量损失32%(Wiseman and Isenhour, 1993)、尼加拉瓜玉米产量损失45% (Hruska and Gladstone, 1988)。草地贪夜蛾具有极强的远距离迁飞和扩散能力(吴秋琳等, 2019),2016年首次报道入侵非洲西部和中部地区,在2017年初扩散到非洲南部地区,对当地玉米产量造成多达21%~53%的损失(Abrahamsetal., 2017)。2019年1月13日确认该虫从缅甸入境我国云南省(杨普云等, 2019)。截至2019年9月11日,草地贪夜蛾已经入侵我国25个省份的1 485个县(市、区),发生面积104万ha,防治面积157万ha(农业农村部种植业管理司种植业快报草地贪夜蛾防控专刊第26期)。由于草地贪夜蛾对玉米等主要农作物的暴食危害性和超强迁飞扩散能力,2017年被国际农业和生物科学中心CABI列为世界十大植物害虫之一(https:∥www.cabi.org/isc/fallarmyworm)。目前草地贪夜蛾已经在北美洲、南美洲、非洲和亚洲四大洲的100多个国家发生为害,并且还在继续蔓延扩张,引起了国际社会广泛关注,其暴食性、强迁飞能力、高繁殖力、对化学农药和Bt杀虫蛋白易产生抗性的特点也为防控带来了严峻挑战。
有机磷和氨基甲酸酯类杀虫剂由于品种多、活性高、杀虫谱广、成本低等特点,最近几十年一直被用于草地贪夜蛾的防治。但是由于长期过量和不当使用,目前草地贪夜蛾已经对这两类杀虫剂产生了明显抗性。早在1979年,美国印第安纳州哈蒙德市的草地贪夜蛾就对氨基甲酸酯类杀虫剂甲萘威产生了41倍的抗性,对有机磷类杀虫剂甲基对硫磷和敌百虫分别产生了113和31倍的抗性(Woodetal., 1981)。1981年美国乔治亚州草地贪夜蛾田间种群对氨基甲酸酯类杀虫剂甲萘威的抗性达90倍,并对有机磷类杀虫剂对硫磷和二嗪农有低水平交互抗性(McCord and Yu, 1987);1989年美国佛罗里达州草地贪夜蛾田间种群对毒死蜱、甲基对硫磷、二嗪农、甲丙硫磷、敌敌畏和马拉硫磷6种有机磷类杀虫剂产生了12~271倍的抗性,同时对灭多威、甲萘威和硫双灭多威3种氨基甲酸酯类杀虫剂产生了14~192倍的抗性(Yuetal., 1991);2002年,佛罗里达北部两块玉米田里采集的草地贪夜蛾幼虫对甲萘威已经产生极高水平抗性,抗性倍数分别为626和1 159倍,对甲基对硫磷分别产生了30和39倍中等水平抗性(Yu and McCord, 2007);同样是在2002年,佛罗里达州斯特拉地区草地贪夜蛾田间种群对甲萘威和甲基对硫磷达到了高抗水平,抗性倍数分别是562和354倍 (Yuetal., 2003);Gutiérrez-Moreno等(2019)报道波多黎各草地贪夜蛾田间种群对毒死蜱抗性倍数是47倍,对灭多威和硫双灭多威的抗性倍数分别达到223和124倍。由这些报道可以看到草地贪夜蛾对有机磷和氨基甲酸酯类杀虫剂产生抗性的速度非常快。
由于草地贪夜蛾化学防治历史比较长,因此多样性的抗药性机制也随之进化,但是主要集中在代谢抗性和靶标抗性两个方面。McCord和Yu (1987)在研究草地贪夜蛾对甲萘威抗性机理时发现,细胞色素多功能氧化酶(P450s)的抑制剂增效醚(piperonyl butoxide, PBO)对甲萘威的增效作用可达15倍,表明P450s可能在抗性产生过程中发挥着重要作用。进一步研究发现,抗性种群对甲萘威氧化降解的速率是敏感种群的5倍,说明抗性种群的氧化代谢作用增强是导致其对甲萘威产生抗性的重要机制。Yu (1992)的研究表明,草地贪夜蛾对多种杀虫剂的广谱抗性除与多功能氧化酶、谷胱甘肽-S-转移酶、水解酶和还原酶等解毒酶活性升高有关外,其乙酰胆碱酯酶对敌敌畏等的敏感性也显著下降。目前已有研究证明乙酰胆碱酯酶基因ace-1的点突变与草地贪夜蛾对有机磷产生抗性有关。如草地贪夜蛾毒死蜱抗性品系的ace-1基因同敏感品系相比,在201A, 227G和290F这3个氨基酸位点发生了点突变 (Carvalhoetal., 2013)。Herrera-Mayorga等(2018)在墨西哥草地贪夜蛾田间种群ace-1中也发现了A201S和F290V突变,但未检测到G227A突变;此外还检测到3个新的突变,但是是否与抗性相关尚不清楚。
由于草地贪夜蛾刚入侵我国,目前我国尚无登记用于防治该虫的杀虫剂,农业农村部根据《农药管理条例》有关规定,推荐了25种用于应急防控草地贪夜蛾的药剂(种植业管理司,2019年6月5日),其中有机磷类杀虫剂乙酰甲胺磷为推荐用药之一。在实际防治中,也同时使用其他有机磷类杀虫剂如毒死蜱和氨基甲酸酯类杀虫剂如甲萘威。鉴于在国外草地贪夜蛾防治中,有机磷和氨基甲酸酯杀虫剂使用时间长、抗性水平高,迫切需要监测入侵我国的草地贪夜蛾田间种群对这两类杀虫剂的抗性基因型及频率。本研究利用DNA检测方法检测了2019年我国12个省市草地贪夜蛾田间种群中有机磷和氨基甲酸酯类杀虫剂靶标基因ace-1的基因型,明确了抗性基因频率。研究结果在分子水平上揭示了入侵我国的草地贪夜蛾携带有高频率对有机磷和氨基甲酸酯类杀虫剂抗性基因,对指导田间合理、科学用药具有重要应用意义。
1 材料与方法
1.1 供试昆虫
草地贪夜蛾田间种群采自我国云南省、广东省、江西省等12个省市区,采集信息详见表1。田间种群个体采集采用随机取样法,每株玉米作为1个采样点,只取1头幼虫,两个间隔采样点至少距离5 m,每个省份采集幼虫至少200头。采集的幼虫活体样本带回实验室,在体视显微镜下根据郭井菲等(2019)描述的草地贪夜蛾形态学特征(头部倒“Y”型纹和第8腹节背面4个黑点)进行鉴定,确认无误后一部分冻存于-80℃冰箱用于本实验抗性基因分子检测,剩余部分个体用于在实验室建立稳定的种群供其他实验使用。

表1 中国12个省份玉米田草地贪夜蛾采集信息Table 1 Sampling data of Spodoptera frugiperda from maize fields in 12 provinces in China
1.2 ace-1基因3个点突变位点的检测
用手术刀切取上述冻存的田间采集的单头幼虫头部或腹部组织50 mg, 采用TIANamp Genomic DNA提取试剂盒(北京天根生化科技有限公司)提取幼虫基因组DNA,幼虫在提取DNA之前先饥饿处理12 h,使其将取食的玉米叶片排泄干净,以避免提取到玉米DNA进而导致PCR的非特异性扩增。其余步骤参照试剂盒说明书进行。提取的幼虫DNA用NanoDrop 1000分光光度计(Thermo Fisher Scientific, Waltham, 美国)测定浓度。敏感和抗毒死蜱品系乙酰胆碱酯酶基因ace-1序列(GenBank序列号: KC435023和KC435024)比对发现有3个突变位点,分别是A201S, G227A和F290V (Carvalhoetal., 2013)。利用Primer Premier 5 (PREMIER Biosoft International, 美国)设计针对这3个突变位点的特异性引物Sfru-ace-1-F: 5′-CCTGATGTACC GGGTAATGCTG-3′和Sfru-ace-1-R: 5′-TAACCTTCC TCCGTATTGGATCC-3′。以单头幼虫DNA为模板,进行PCR扩增。PCR反应体系(25 μL): TaKaRaExTaq酶(5 U/μL) 0.125 μL, 10×ExTaq Buffer 2.5 μL, dNTP Mixture (各2.5 mmol/L)2 μL, 正反向引物(10 μmol/L)各2.5 μL, DNA模板(200 ng/μL)2.5 μL,无菌水12.875 μL。反应程序: 94℃预变性3 min; 94℃ 1 min, 56℃ 1 min, 72℃ 45 s, 36个循环反应; 最后72℃延伸10 min。PCR产物由1.2%琼脂糖凝胶电泳检测,条带正确的PCR产物用基因特异性引物进行3730XL(Applied Biosystems, 美国) DNA双向测序。
1.3 数据分析
使用Chromas 2.31软件对测序峰图进行分析,核对ace-1编码蛋白在A201S, G227A和F290V位点对应核苷酸是否发生点突变。 Excel记录统计每个幼虫个体的基因型,计算每个种群的抗性基因频率。 抗性基因频率%=[(抗性纯合子个体数/检测总数)+(抗性杂合子个体数/检测总数/2)]×100%。
2 结果
2.1 草地贪夜蛾乙酰胆碱酯酶基因ace-1的3个突变位点分析
草地贪夜蛾对毒死蜱敏感和抗性品系ace-1基因序列(GenBank序列号: KC435023和KC435024)(Carvalhoetal., 2013)比对发现,在抗性品系中发生了3个点突变,分别是GCG突变成TCG(下划线示发生突变的核苷酸,下同),对应氨基酸由丙氨酸突变成丝氨酸(A201S, 以下简称201);GGA突变成GCA,对应氨基酸由甘氨酸突变成丙氨酸(G227A,以下简称227);TTT突变成GTT,对应氨基酸由苯丙氨酸突变成缬氨酸(F290V,以下简称290)。
2.2 草地贪夜蛾乙酰胆碱酯酶基因ace-1基因型和抗性基因频率分析
通过特异性引物Sfru-ace-1-F和Sfru-ace-1-R扩增得到长度为500 bp的ace-1基因片段(图1: A)。通过PCR扩增和DNA测序一共检测了我国12个省份的田间种群个体589头,通过峰图分析发现,ace-1基因跟有机磷抗性相关的3个位点在589头个体之间检测出不同的基因型,201位点有2种基因型:敏感纯合型SS和抗性杂合型RS;227位点在检测样本中均为敏感纯合型SS;290位点检测出3种基因型:敏感纯合型SS、抗性杂合型RS和抗性纯合型RR(图1: B)。
各省份田间种群中,201位点63.9%~88.6%的个体为敏感纯合子,11.4%~36.1%的个体为抗性杂合子,无抗性纯合子,抗性基因频率最低为江苏省5.7%,最高为四川省18.0%, 12省份的平均抗性基因频率为11.4%。227位点100%的个体为敏感纯合子,抗性基因频率为0。290位点3.3%~57.6%的个体为抗性纯合子,40.7%~77.3%的个体为抗性杂合子,抗性基因频率最低为重庆市39.2%,最高为江西省78.0%,12个省份的平均抗性基因频率为56.9%。290位点的抗性基因频率显著高于201和227位点(表2)。
3 讨论
有机磷类和氨基甲酸酯类杀虫剂用于防治草地贪夜蛾已经有30多年的历史,由于长期不合理用药,导致草地贪夜蛾对多种有机磷和氨基甲酸酯类杀虫剂产生了抗药性。乙酰胆碱酯酶(AchE, EC 3.1.1.7)点突变导致的对有机磷和氨基甲酸酯类杀虫剂的不敏感性是导致抗性产生的重要原因(Walshetal., 2001)。在不同种类蚊子中已报道两个乙酰胆碱酯酶AChE1和AChE2,分别由ace-1和ace-2基因编码,AChE1和AChE2的突变都会导致昆虫对有机磷和氨基甲酸酯类杀虫剂产生抗性(Chenetal., 2007; Kim and Lee, 2013; Liu, 2015; Wuetal., 2015)。巴西种群草地贪夜蛾毒死蜱抗性品系ace-1基因同敏感品系相比在201, 227和290这3个氨基酸位点发生了点突变,其中201位点的抗性基因最低,为17.5%,仅检测到1头纯合突变个体;227和290位点的抗性基因频率较高,分别为67.5%和32.5%(Carvalhoetal., 2013)。墨西哥草地贪夜蛾田间种群ace-1基因检测结果显示,在田间采集的40头个体中201位点发生了12.5%的纯合抗性突变;227位点没有发现杂合或纯合突变个体,100%为纯合敏感基因型SS;290位点发生了50%的突变,其中25%纯合突变,25%杂合突变(Herrera-Mayorgaetal., 2018)。本研究在检测的我国12省份589头个体中,201位点有137头个体(23.3%)为抗性杂合个体,没有发现抗性纯合个体。同墨西哥种群检测结果一样,在227位点100%为敏感纯合个体,没有发现杂合或纯合抗性个体,表明入侵我国的草地贪夜蛾种群本就不携带这种突变,而入侵之后也没有发生该位点的突变。在我国草地贪夜蛾589头个体中,290位点的抗性基因频率最高,携带抗性基因的个体数量达到523头(占样本总数的88.8%),其中373头个体(63.3%)携带抗性杂合RS基因,150头个体(25.5%)携带抗性纯合RR基因(表2)。巴西、墨西哥和我国草地贪夜蛾种群ace-1基因型和抗性基因频率的差异可能是由于个体接触的杀虫剂种类和选择压不同导致的。但是值得注意的是,从检测数量上来说,本研究的589头个体要远远多于巴西种群的20头和墨西哥的40头,所得结论能够更加真实反映目前我国不同省份草地贪夜蛾田间种群ace-1的基因型以及抗性基因频率。
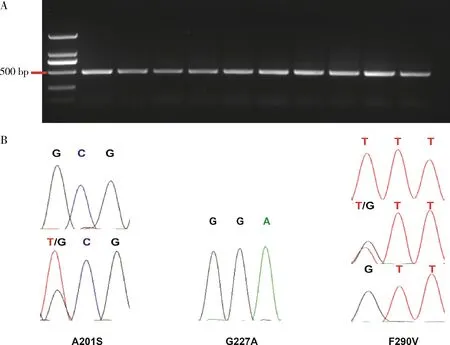
图1 草地贪夜蛾ace-1基因突变位点分析Fig. 1 Analysis of ace-1 gene mutation sites in Spodoptera frugiperdaA: 用特异性引物随机克隆的10个个体ace-1 PCR产物PCR products of ace-1 gene of 10 randomly selected individuals using gene specific primers; B: 草地贪夜蛾ace-1基因突变位点分型图Genotyping of ace-1 gene mutation sites in S. frugiperda.
另外,除了A201S, G227A和F290V 3个突变位点,近期在墨西哥种群中报道了3个新的突变位点g-396 G/A, g-498 A/G, g-768 C/G (Herrera-Mayorgaetal., 2018),但是经分析这不是3个新的突变位点,而是正常的核苷酸多态性现象,因为这3个核苷酸位点的突变,并没有引起对应编码的氨基酸发生改变(g-396 G/A位点CCG突变成CCA,但是编码的氨基酸不变,都是脯氨酸;g-498 A/G位点GAA突变成GAG,但是编码的氨基酸不变,都是谷氨酸;g-768 C/G位点GTC突变成GTG,但是编码的氨基酸不变,都是缬氨酸)。本研究对我国12个种群589头个体的ace-1测序峰图分析没有发现新的突变位点。Carvalho等(2013)在检测的20头毒死蜱抗性品系个体中有8头在227和290这2个位点以抗性杂合基因型形式共同出现,另有1头在201和227位点以抗性纯合型共同出现。本研究检测结果显示,589头个体在227位点均为敏感纯合型,同Herrera-Mayorga等(2018)对墨西哥种群的检测结果相一致;有137头在201位点为抗性杂合型(表2),其中有127头(92.7%)在201和290位点同时以抗性杂合型出现,推测在201位点为抗性杂合基因型的个体,其在290位点通常也会产生杂合突变;有150头在290位点为抗性纯合型(表2),其中有149头(99.3%)在201位点同时为敏感纯合型,推测在290位点为抗性纯合基因型的个体,其在201位点通常不会发生杂合或纯合突变。上述3个位点的规律性突变特点可以作为我国草地贪夜蛾有机磷和氨基甲酸酯靶标基因ace-1突变快速检测的参考依据,具有重要的现实意义。

根据迁飞习性、气象学数据和迁飞轨迹模拟发现,目前草地贪夜蛾是通过东、西两条路径入侵我国的,东线主要从越南、老挝、泰国进入广东广西两省,然后依靠强劲的西南风向北入侵长江流域和华北玉米主产区省份;西线主要从缅甸进入云南,然后经贵州进入重庆、四川和陕西(Lietal., 2019)。东西两条线草地贪夜蛾迁飞速度不同,加上各省份用药种类和频率也有不同,最终会导致抗性产生速度不同。在西线上的云南、贵州、重庆和四川4个省份,201位点的抗性基因频率呈现出依次升高的趋势,而290位点的抗性基因频率在西线5省上无明显差异(表2),这可能是这两个位点在药剂选择压下的突变速率不同导致。在东线上的广东、江西、福建、江苏、湖南和安徽6个省份,201和290这两个位点的抗性基因频率无明显规律特征(表2),可能跟草地贪夜蛾借助强劲的西南风快速随机北迁,药剂选择压不强相关。
化学杀虫剂作为害虫防治的“压舱石”和“杀手锏”,对2019年成功防治草地贪夜蛾发挥了至关重要的作用。由于我国转基因玉米还没有商业化种植,所以未来几年防治草地贪夜蛾化学杀虫剂还将会扮演主要角色。但是我们应该清楚地看到,一方面,入侵我国的草地贪夜蛾种群本身就携带高频率的抗性基因;另一方面,我国不同省份之间用药种类和用药频率有很大差异,害虫产生抗药性的速度和水平也会不同,所以根据我国不同省份种植模式、用药种类和频率,展开不同省份草地贪夜蛾田间种群抗药性监测和治理工作就显得尤为重要(李永平等, 2019; 吴益东等, 2019)。本研究通过检测全国12省份的草地贪夜蛾589头个体ace-1基因型和抗性基因频率,发现跟有机磷和氨基甲酸酯类杀虫剂抗性相关的3个突变位点(第201, 227和290位氨基酸)中有两个(201和290位点)存在不同频率的突变,尤其是290位点的突变频率最高,达到了57.1%。表明入侵中国的草地贪夜蛾携带高频率的抗性基因,同时进入我国后再经过杀虫剂的多轮选择作用,推测已经对有机磷和氨基甲酸酯类杀虫剂产生了较高的抗性。在实际田间防治中,应该不用或尽量少用有机磷和氨基甲酸酯类杀虫剂。有报道表明,甲氨基阿维菌素苯甲酸盐、乙基多杀菌素、氯虫苯甲酰胺、甲维盐等对草地贪夜蛾均有非常强的杀虫活性(王勇庆等, 2019; 尹艳琼等, 2019; 赵胜园等, 2019),在田间防治中可以优先使用,但同时也要加强抗性监测,以检测草地贪夜蛾对这些药剂是否产生抗药性。
最后需要指出的是,除了靶标基因发生点突变,解毒酶的活性增强也是草地贪夜蛾对有机磷和氨基甲酸酯类杀虫剂产生抗性的重要原因。在巴西草地贪夜蛾毒死蜱抗性品系中发现P450s, GSTs和CXEs基因表达量上升(Carvalhoetal., 2013);对草地贪夜蛾基因组分析发现,P450s和GSTs两大解毒代谢酶基因家族发生了严重扩张,这可能是草地贪夜蛾高杂食性和易对化学农药产生抗性的原因之一(Liuetal., 2019)。因此,对入侵我国的草地贪夜蛾对有机磷和氨基甲酸酯类杀虫剂的代谢抗性机制也有待进一步加强研究。
猜你喜欢
云南农业(2021年11期)2021-11-12
毛纺科技(2021年8期)2021-10-14
食品安全导刊(2021年21期)2021-08-30
食品安全导刊(2021年20期)2021-08-30
今日农业(2020年13期)2020-12-15
农药科学与管理(2019年7期)2019-11-29
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
湖北农业科学(2016年21期)2017-03-18
中国社区医师(2006年7期)2006-04-19
