
草地贪夜蛾热激蛋白基因SfHsp90的克隆及在高低温和UV-A胁迫下的表达分析
2020-07-08 00:53孟建玉杨昌利胡朝兴张长禹
昆虫学报 2020年5期
周 吕, 孟建玉, 杨昌利, 李 锦, 胡朝兴, 张长禹,*
(1. 贵州大学昆虫研究所, 贵州省山地农业病虫害重点实验室, 贵阳 550025; 2. 贵州省烟草科学研究院, 贵阳 550081)
热激蛋白(heat shock proteins, Hsps)又被称为热休克蛋白,是一类广泛存在于各种生物体内并在进化上高度保守的特殊蛋白质(Zhao and Jones, 2012; 司风玲等, 2016)。自然界中,热激蛋白可在多种生物胁迫(如真菌、细菌、病毒、昆虫等)和非生物胁迫(如UV、重金属、高温、低温、水分、药剂、盐浓度等)下诱导表达(马文静和马纪, 2012)。热激蛋白主要功能是作为分子伴侣参与到蛋白质的合成、折叠、组装过程中,影响细胞的分泌和调节活动,是生物适应环境胁迫的重要适应机制之一(Parsell and Lindquist, 1993)。根据热激蛋白的分子量,可将热激蛋白分为Hsp90, Hsp70, Hsp60, Hsp40和小分子热激蛋白(Feder and Hofmann, 1999; Sørensenetal., 2003; Gkouvitsasetal., 2009)。Hsp90是一类正常细胞中不可缺少的蛋白质,广泛存在于原核生物和真核生物中,在昆虫生长发育和对极端环境的适应中发挥着重要作用(Rutherfordetal., 2007; 贾栋, 2015; 姜建军等, 2019)。如烟蚜MyzuspersicaeHsp90基因在UV-B胁迫下的表达量显著上升,并随着照射时间的延长呈现出先上升后下降的趋势(苏丽等, 2018)。高温对甜菜夜蛾Spodopteraexigua幼虫体内Hsp90基因表达有明显诱导作用,表达量随温度增加呈上升趋势(翟会芳等, 2010)。
近年来,由于环境变化,大气中的臭氧含量减少,导致地球表面的紫外线辐射越来越强(Xu and Sullivan, 2010)。根据波长,可将紫外线分为UV-A, UV-B和UV-C。其中UV-A辐射可以影响生物的生长发育和行为选择(Verdagueretal., 2016; Zhouetal., 2018)。Zhang等(2011)研究发现,UV-A照射会使棉铃虫Helicoverpaarmigera成虫的寿命缩短而生殖力提升。UV-A照射还能诱导棉铃虫雌虫体内JNK基因的表达(刘小飞等, 2019)。温度也是影响昆虫生理生化的重要因素。在高温处理下,昆虫会出现生长速率下降、滞育、畸形和无法交配产卵等现象(Butleretal., 1985; 马春森和陈瑞鹿, 1993; Williamsetal., 2003; 罗举等, 2005)。低温会影响昆虫的存活率、寿命和相关基因的表达(马春森等, 2005; 孔雪华等, 2012; 李耀华等, 2017)。沙葱萤叶甲Galerucadaurica2龄幼虫经过-10, -5和0℃的低温胁迫1 h,25℃恢复30 min后,其表皮蛋白基因GdAbd表达显著上调(单艳敏等, 2019)。
草地贪夜蛾Spodopterafrugiperda属鳞翅目(Lepidoptera)夜蛾科(Noctuidae)灰翅夜蛾属Spodoptera,又名秋黏虫,是一种广泛分布于美洲大陆热带和亚热带地区的杂食性昆虫(Todd and Poole, 1980)。幼虫可取食玉米、水稻、高粱、棉花和蔬菜等多种农作物(Montezanoetal., 2018)。草地贪夜蛾成虫具有极强的迁飞能力,自2019年1月在我国云南被发现,至6月已经扩散至我国南方18个省(Goergenetal., 2016; Feldmannetal., 2019; 唐璞等, 2019)。目前,国内有关草地贪夜蛾响应环境胁迫的分子机制研究较少。本研究通过克隆草地贪夜蛾体内Hsp90基因,分析其序列特征和表达谱,检测其在不同龄期和成虫不同组织中的表达,研究其在UV-A、高温和低温胁迫下的差异表达,揭示Hsp90基因在响应环境胁迫中的作用,探索草地贪夜蛾响应环境胁迫的分子机制,为探究草地贪夜蛾适应环境胁迫的机理,制定科学的防治策略提供参考。
1 材料与方法
1.1 试虫
本实验所用草地贪夜蛾于2019年7月在贵州省安顺市普定县野外玉米田中采集获得。成虫由野外采集的幼虫羽化而成,幼虫为实验室玉米叶饲养后代。饲养条件:温度25±1℃,相对湿度70%±5%,光周期16L∶8D。
1.2 样本处理
选取草地贪夜蛾卵、1-6龄幼虫、蛹、成虫作为不同发育阶段样本。选取羽化后3 d雌雄成虫的头(去除触角和复眼)、胸、腹、触角、复眼、足、中肠、翅、精巢(雄虫)和卵巢(雌虫)等作为组织样本。取羽化后3 d的雌雄成虫,设置4℃和36℃的温度处理,处理时间为0(CK), 30, 60, 90, 120和150 min,作为不同温度处理样本。选取羽化后3 d的雌雄成虫,暗适应2 h后,以UV-A(波长315~400 nm)紫外灯为光源,对草地贪夜蛾雌雄成虫进行照射处理,照射强度为300 μW/cm2,处理时间为0(CK), 30, 60, 90, 120和150 min,照射期间温湿度和正常饲养条件一致,作为UV-A处理样本。每个处理设置3个生物学重复,各发育阶段每重复样本数分别为:卵50粒,1龄幼虫20头,2-3龄幼虫10头,4-6龄幼虫、蛹和成虫均为8头;组织样本每个重复解剖8头成虫。
1.3 总RNA提取和cDNA合成
使用Eastep® Super Total RNA Extraction Kit试剂盒提取草地贪夜蛾总RNA。用1%琼脂糖凝胶电泳检测RNA的质量,使用NanoPhotometerTMP-Class分光光度仪检测其浓度,按照HiFiScript cDNA Synthesis Kit说明书完成cDNA第1链的合成。
1.4 草地贪夜蛾Hsp90基因的克隆
通过对已知同源昆虫斜纹夜蛾Spodopteralitura的Hsp90基因(GenBank登录号: AF254880.1)序列进行比对,在保守区设计1对简并引物Hsp90-F/Hsp90-R(表1),以1.3节合成的cDNA第1链为模板扩增草地贪夜蛾Hsp90基因的中间片段。PCR反应体系(25 μL): TaKaRa Ex Primer TaqTM酶(1.25 μmol/25 μL)12.5 μL, cDNA模板1.5 μL, 正反向引物(10 mmol/L)各1.5 μL, DEPC水8 μL。反应条件: 95℃ 3 min, 95℃ 30 s, 55℃ 30 s, 72℃ 1 min, 35个循环;72℃ 5 min。用1%琼脂糖凝胶电泳检测PCR产物,使用SanPrep柱式PCR产物纯化试剂盒回收目的片段,连接到pMDTM19-T载体并转化到大肠杆菌EscherichiacoliDH5α感受态细胞,挑取单一菌落经PCR检测正确后进行扩大培养,菌液送往上海生工生物有限公司测序。
在获得的目的基因片段的基础上继续向两端设计引物Hsp90-F2/Hsp90-R2和Hsp90-F3/Hsp90-R3(表1),以cDNA第1链为模板,经PCR反应后,扩增出草地贪夜蛾编码区序列。对所得产物进行纯化回收,送往上海生工生物工程有限公司测序。

表1 引物信息Table 1 Primer information
1.5 草地贪夜蛾Hsp90基因的序列分析
利用DNAMAN6.0对所得基因片段进行拼接,使用NCBI(https:∥blast. Ncbi.nlm.nih.gov/Blast.cgi)中的Blast工具进行氨基酸的同源性分析;利用ORFfinder (http:∥www.ncbi.nlm.nih.gov/projects/gorf/)分析草地贪夜蛾Hsp90基因的开放阅读框;使用ExPASy ProtParam Tool (https:∥web.expasy.org/cgi-bin/protparam/protparam)、KinasePhos和ExPASy Tools中的NetNGlyc预测蛋白质的理化性质;以脊索动物门青鱼Mylopharyngodonpiceus的Hsp90(GenBank登录号:AVO65120.1)为外群,利用NCBI数据库进行基因比对,使用MEGA6.0采用邻接(neighbor-joining)法1 000次构建草地贪夜蛾和其他昆虫Hsp90的系统发育树;应用ExPASy ScanProsite (https:∥prosite.expasy.org/ scanprosite/)进行家族签名序列分析。
1.6 草地贪夜蛾Hsp90基因的表达分析
取1.2节草地贪夜蛾不同处理材料,采用Eastep® Super Total RNA Extraction Kit试剂盒分别提取总RNA,用反转录试剂盒(HiFiScript cDNA Synthesis Kit)反转录成cDNA,采用RT-qPCR分析草地贪夜蛾Hsp90基因在草地贪夜蛾不同发育阶段和成虫组织中的表达量差异,以及其在不同时长UV-A胁迫和不同温度胁迫下草地贪夜蛾成虫中表达量的变化。根据草地贪夜蛾Hsp90基因的全长序列设计1对定量引物qHsp90-F/qHsp90-R;选用核糖体蛋白L27(ribosomal protein L27, RPL27)基因作为内参基因,Sf-RPL27-qPCR-F/Sf-RPL27-qPCR-R作为内参基因引物(Karamipouretal., 2018)(表1),采用SYBR Green Supemix 染料(TaKaRa公司)法进行RT-qPCR,反应体系(20 μL): cDNA模板2 μL, 2×iTaqTMSYBR Green Supermix 10 μL, 上下游引物(10 μmol/L)各1 μL, ddH2O 6 μL。反应条件: 95℃ 3 min; 95℃ 30 s, 58℃ 30 s, 进行39个循环; 65℃ 5 s。
1.7 数据分析
利用2-△△CT法(Livak and Schmittgen, 2001)对草地贪夜蛾Hsp90基因的表达水平进行相对定量分析。实验数据用SPASS21.0软件中的Duncan氏多重比较对数据进行统计分析。
2 结果
2.1 草地贪夜蛾Hsp90基因克隆与序列分析
以草地贪夜蛾的cDNA为模板,经克隆得到2 323 bp基因序列,命名为SfHsp90(GenBank登录号: MN832694),经NCBI分析可知开放阅读框长2 154 bp,编码717个氨基酸(图1)。草地贪夜蛾Hsp90 C末端序列为EEVD(图2),表明该蛋白具有细胞质型热激蛋白氨基酸特征,属于胞质热激蛋白;编码蛋白相对分子量为82.52 kD;等电点(pI)为5.01;带负电荷的残基总数(Asp+Glu)为144个,带正电荷的残基总数(Arg+Lys)为107个;分子式为C3634H5798N968O1161S28;N末端是M(Met),推测半衰期为30 h,稳定性系数为39.23,说明该蛋白质性质稳定。总平均疏水指数为-0.679,提示其为亲水性蛋白质。

图1 草地贪夜蛾热激蛋白基因SfHsp90的核苷酸序列以及推断的氨基酸序列Fig. 1 Nucleotide and deduced amino acid sequences of heat shock protein gene SfHsp90 from Spodoptera frugiperda单下划线代表起始密码子ATG, 星号代表终止密码子TAA, 框内为Hsp90的家族签名序列,双下划线是糖基化位点,阴影部分标注的是磷酸化位点,单波浪下划线是C端共识序列EEVD。The single underline represents the initiation codon ATG, the asterisk represents the termination codon TAA, the family signature sequence of Hsp90 is in the box, the double underline is the glycosylation site, the shaded part is the phosphorylation site, and the single wavy underline is the C-terminal consensus sequence EEVD.

图2 草地贪夜蛾与其他物种Hsp90的多序列比对Fig. 2 Multiple sequence alignment of Hsp90 from Spodoptera frugiperda and other speciesHsp90来源物种GenBank登录号Origin species of Hsp90 and their GenBank accession numbers: SfHsp90: 草地贪夜蛾Spodoptera frugiperda (MN832694); SlHsp90: 斜纹夜蛾Spodoptera litura (XP_022834656.1); LsHsp90: 草地螟Loxostege sticticalis (ABW87791.1); HaHsp90: 异色瓢虫Harmonia axyridis (AWV96614.1); MpHsp90: 青鱼Mylopharyngodon piceus (AVO65120.1).
2.2 草地贪夜蛾Hsp90系统进化分析

图3 邻接法构建的基于Hsp90氨基酸序列的系统发育树Fig. 3 Phylogenetic tree of Hsp90 constructed by neighbor-joining method based on amino acid sequence利用Bootstrap进行1 000次计算以检验进化树分支置信度,分支上数值表示置信度。Bootstrap was used for 1 000 calculations to test the confidence of the branch of the evolutionary tree. The value on the branch indicates the confidence.
在NCBI数据库中搜索鳞翅目、鞘翅目、双翅目和半翅目22种昆虫和外群生物青鱼Mylopharyngodonpiceus的Hsp90氨基酸序列,与推导的草地贪夜蛾SfHsp90氨基酸序列采用邻接法构建系统发育树(图3)。结果表明,系统发育树分成2个大的分支,其中昆虫纲23种昆虫聚为一支,外群生物青鱼单独聚为一支,表明Hsp90的高度保守性;昆虫纲中,鳞翅目、鞘翅目、双翅目和半翅目昆虫分别聚为一支,与传统昆虫分类学结果一致。从目以下的分类阶元来看,草地贪夜蛾与斜纹夜蛾Spodopteralitura、棉铃虫Helicoverpaarmigera、苜蓿夜蛾Heliothisviriplaca、大螟Sesamiainferens、二化螟Chilosuppressalis、草地螟Loxostegesticticalis、夏威夷红蛱蝶Vanessatameamea、落叶松毛虫Dendrolimussuperans聚为一支,说明其亲缘关系较近,其中草地贪夜蛾与斜纹夜蛾亲缘关系最近,同其他几个目昆虫的亲缘关系较远,和鱼纲中的青鱼亲缘关系最远。
2.3 草地贪夜蛾SfHsp90基因的表达分析
SfHsp90基因在不同发育阶段的表达量存在显著的差异(P<0.05), 在1龄幼虫中的表达量最高,其次是在卵中的表达量,在2龄和3龄幼虫期也有较高的表达量;4-6龄幼虫中的表达量较低,6龄幼虫表达量最低(图4)。SfHsp90在成虫不同组织中的表达量也存在显著差异(P<0.05),在雄成虫头部(去除触角和复眼)、触角、复眼、中肠和精巢中表达量较高,在其余部位表达量较低;在雌成虫头部(去除触角和复眼)、触角、复眼和卵巢中有较高的表达,在其他部位的表达量较低(图5)。

图4 草地贪夜蛾SfHsp90在不同发育阶段的相对表达水平Fig. 4 Relative expression levels of SfHsp90 in different developmental stages of Spodoptera frugiperdaE: 卵Egg; L1-L6: 分别为1-6龄幼虫1st-6th instar larva, respectively; P: 蛹Pupa; M: 雄成虫Male adult. 数据为平均值±标准差,柱上不同小写字母表示不同发育阶段间的相对表达量存在显著性差异(P<0.05, Duncan氏多重比较)。Data are mean±SD. Different lowercase letters above bars indicate significant difference in the expression level between different developmental stages (P<0.05, Duncan’s multiple range test).

图5 草地贪夜蛾雌雄成虫不同组织中SfHsp90的相对表达水平Fig. 5 Relative expression levels of SfHsp90 in different tissues of male and female adults of Spodoptera frugiperdaHD: 去除触角和复眼的头部Head with antennae and compound eyes removed; TX: 胸部Thorax; AN: 腹部Abdomen; AA: 触角Antenna; CE: 复眼Compound eye; FT: 足Leg; WG: 翅Wing; MT: 中肠Midgut; TS: 精巢Testis; OY: 卵巢Ovary. 数据为平均值±标准差,柱上不同小写字母表示不同组织间的相对表达量存在显著性差异(P<0.05, Duncan氏多重比较)。Data are mean±SD. Different lowercase letters above bars indicate significant difference in the relative expression level between different tissues (P<0.05, Duncan’s multiple range test).
如图6所示,36℃处理下,除雌虫30 min处理外,雌雄成虫SfHsp90表达量均显著高于对照(0 min) (P<0.05);雄成虫的SfHsp90表达量随着处理时间的增加呈现出先升高后降低的趋势,在处理时间为60 min时达到最高值,在90, 120和150 min时表达量逐渐下降;雌成虫的SfHsp90表达量随着处理时间的增加而升高,在处理时间为150 min时达到最高值。

图6 不同时长36℃高温处理下草地贪夜蛾雌雄成虫SfHsp90的相对表达量Fig. 6 Relative expression levels of SfHsp90 in male and female adults of Spodoptera frugiperdaexposed to high temperature of 36℃ for different time数据为平均值±标准差,柱上不同小写字母表示不同处理时间的相对表达量存在显著性差异(P<0.05, Duncan氏多重比较)。 Data are mean±SD. Different lowercase letters above bars indicate significant difference in the relative expression level between different exposure time (P<0.05, Duncan’s multiple range test). 图7和8同The same for Figs. 7 and 8.
如图7所示,在4℃的低温处理下,SfHsp90的表达量均显著高于对照(0 min)(P<0.05),其中,雄成虫中的表达量呈现先上升后下降的趋势,在30 min处理时达到最高值,随后表达量逐渐下降;雌成虫中的表达量随着处理时间的增加呈逐渐升高的趋势,在处理时间到达150 min时达到最高值。

图7 不同时长4℃低温处理下草地贪夜蛾雌雄成虫SfHsp90的相对表达量Fig. 7 Relative expression levels of SfHsp90 in male and female adults of Spodoptera frugiperda exposedto low temperature of 4℃ for different time
如图8所示,不同时长UV-A处理下,SfHsp90表达量与对照均有显著差异(P<0.05),且随着处理时间的增加,雄成虫体内SfHsp90的表达量呈现先升高后降低趋势,在处理时间为90 min的时候达到最高值;雌成虫体内的表达量也会随着处理时间的增加呈现先升高后降低的趋势,在处理时间为60 min时为最高值。
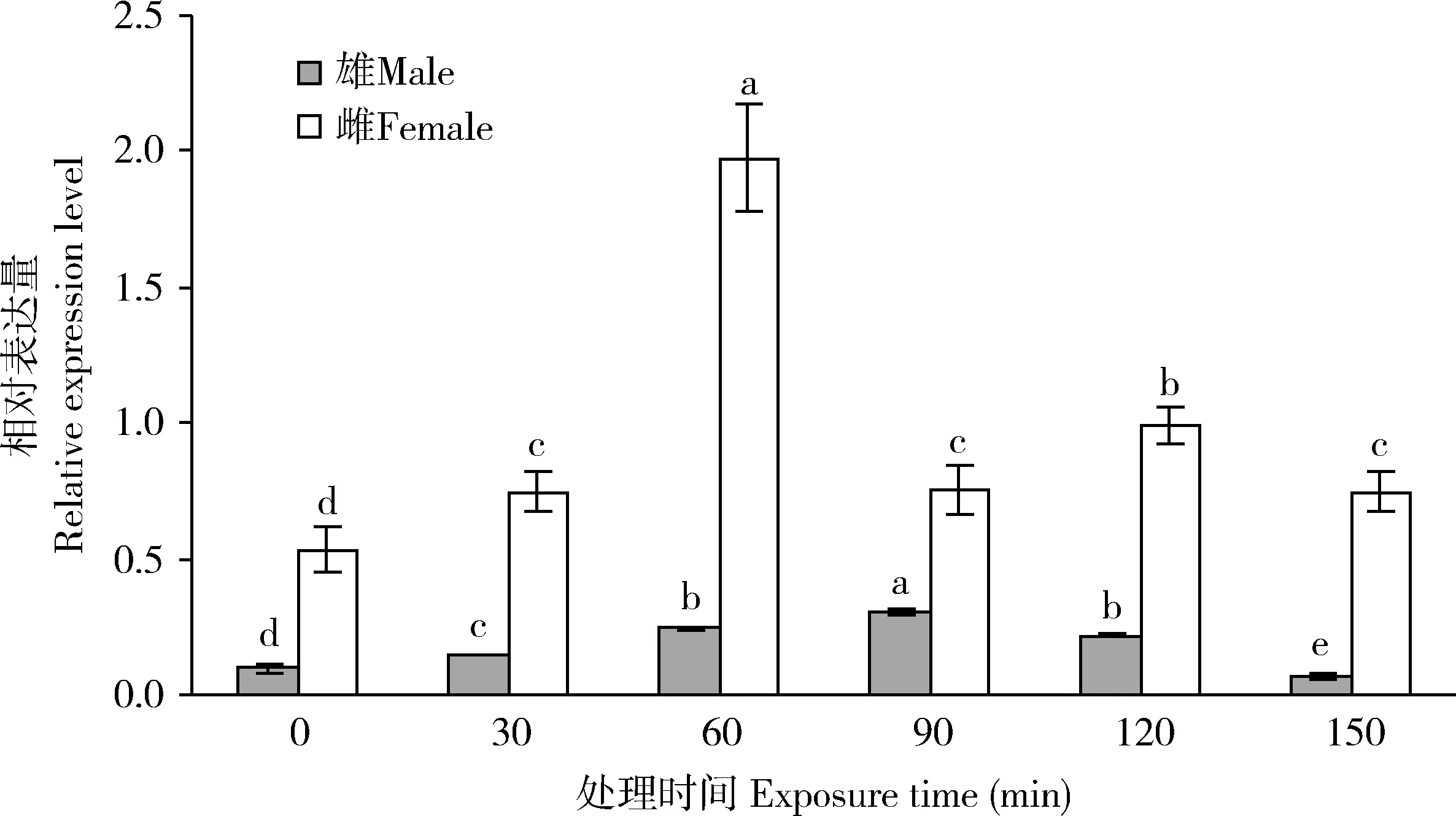
图8 不同时长UV-A处理下草地贪夜蛾雌雄成虫SfHsp90的相对表达量Fig. 8 Relative expression levels of SfHsp90 in male and female adults of Spodoptera frugiperda exposedto UV-A for different time
3 讨论
本研究通过斜纹夜蛾Hsp90设计简并引物,通过RT-PCR技术成功克隆了草地贪夜蛾Hsp90基因SfHsp90,该基因开放阅读框长2 154 bp,编码717个氨基酸。经氨基酸序列分析可知,该基因具有1个家族签名序列,4个糖基化位点和11个磷酸化位点。使用DNAMAN6.0进行氨基酸同源性分析可知,SfHsp90与其他物种的Hsp90具有较高的同源性,其中同源性最高的是斜纹夜蛾Hsp90;系统发育分析结果表明,亲缘关系相近的同一个目的昆虫聚为一支,表明Hsp90在进化中相对保守,这与绿盲蝽Apolyguslucorμm和烟蚜等昆虫的研究结果(Sunetal., 2014; 苏丽等, 2018)一致。
热激蛋白Hsp90是一种广泛存在于真核生物细胞中的蛋白质,当生物受到外界环境的胁迫时,其表达量会出现显著变化(Buchner, 1996; Whitesell and Lindquist, 2005)。通过与多种酶和蛋白之间的相互作用,可有效稳定蛋白的失活,在昆虫生长发育、热胁迫和适应极端温度时发挥着重要作用(Jakob, 1995a, 1995b; Buchner, 1996; Fengetal., 2010)。本实验发现,SfHsp90在不同发育阶段中的表达量差异显著,1龄幼虫时表达量最高,幼虫期时随着发育时间增加表达量逐渐减少,蛹期和成虫期表达量逐渐上升(图4)。这与张敏等(2009)对中红侧沟茧蜂Microplitismediator的研究结果一致。然而,Sonoda等(2006)研究发现,小菜蛾PlutellaxylostellaHsp90在整个发育阶段表达量都没有显著的变化;斜纹夜蛾Hsp90在成虫中的表达量最高(Shuetal., 2011)。由此可见,不同物种在发育阶段的表达具有不同的规律,说明Hsp90在不同物种间的表达有着特异性。不同组织间的基因表达水平亦有差异,本研究发现,SfHsp90在不同组织中均有表达,且不同组织间表达量差异显著(图5),这与林亚秋等(2009)对草鱼Ctenopharyngodonidellus的研究结果相一致。SfHsp90在头(去除触角和复眼)、触角、复眼中的表达量显著高于其他组织(图5),这可能是由于头部是昆虫感觉的中心,触角和复眼是昆虫接受外界信息的重要器官,是时刻感应着外界环境变化的敏感部位,所以导致了Hsp90的高表达(魏国树等, 2002; 那杰等, 2008)。
本研究发现,草地贪夜蛾雌雄成虫在36℃高温和4℃低温处理后,体内的SfHsp90表达量显著提高(图6和7)。这说明Hsp90可响应环境温度胁迫,帮助昆虫适应外界条件的变化(杨静等, 2017)。在两种温度胁迫下,随着处理时间延长,草地贪夜蛾雄成虫的SfHsp90表达量都呈现出先上升后下降的趋势,36℃处理在60 min时达到最高值,4℃处理在30 min时达到最高值,随后表达量都逐渐下降(图6和7)。这可能是因为Hsp90的保护作用有其阈值存在,在环境胁迫超过一定限度时,Hsp90的表达量会下降。这与Cheng等(2016)发现麦红吸浆虫Sitodiplosismosellana幼虫Hsp90在高低温胁迫时其表达量都呈现先上升后下降的结果相符。杨静等(2017)对粘虫的研究也表明,随着处理时间的延长,高温和低温处理下的Hsp90表达量也会呈现先升高后降低的趋势。草地贪夜蛾雌成虫SfHsp90表达量变化趋势与雄成虫不同;两个温度处理下,雌成虫SfHsp90的表达量都显著高于对照(30 min除外),且随着处理时间的增加,表达量逐渐增加,在150 min时达到最高值(图6和7)。这表明草地贪夜蛾雌成虫在遭遇到温度胁迫时,其体内SfHsp90修复细胞的时间阈值在150 min或更长,远远大于雄成虫,说明雌成虫对环境中的温度胁迫具有更强的适应能力。郅伦山(2005)研究中华通草蛉Chrysopasinica发现雌虫的越冬存活率显著高于雄虫,与本研究结果相符。Renault等(2002)的研究也指出,性别会影响昆虫的越冬存活率。
作为太阳紫外线辐射的主要成分之一,UV-A辐射影响昆虫体内多种基因的表达(Alietal., 2017; 刘小飞等, 2019)。本研究发现,不同时间(雄成虫处理150 min除外)UV-A处理下SfHsp90的表达量都显著提高(图8),表明UV-A照射能诱导Hsp90的表达。这与昆虫面对温度和药剂胁迫时的研究结果 (蒋健等, 2013; 姜建军等, 2019)相符。草地贪夜蛾雌雄成虫SfHsp90表达量随着处理时间的延长呈现先上升后下降的趋势,在雄成虫中照射90 min时表达量最高,在雌成虫中照射60 min达到最高值(图8)。这可能是因为UV-A的胁迫造成了草地贪夜蛾细胞损伤,导致了SfHsp90的表达量上调,以修复细胞损伤、抵御UV-A辐射(Jurkiewicz and Buettner, 1994)。随后,长时间的照射导致SfHsp90表达量出现下降趋势(图8),可能是因为长时间的UV-A胁迫对昆虫造成的损伤超过Hsp90保护作用的限度,这与Didomenico等(1982)研究Hsp70得出的结果相似。此外,草地贪夜蛾在不同时长的UV-A照射下,雌成虫SfHsp90的表达量均高于雄成虫的(图8),推测草地贪夜蛾雌成虫对UV-A辐射的抵抗能力可能高于雄成虫。
近年来,大气臭氧层被破坏,随之而来的太阳辐射紫外线增强,全球气候逐渐变暖等问题不可避免地加重了自然界中生物的生存压力。草地贪夜蛾作为一种入侵害虫,其如何抵抗环境胁迫的相关机理研究仍有欠缺。本研究初步阐明了SfHsp90在响应高低温和UV-A辐射胁迫的表达变化,为进一步探索昆虫响应环境胁迫时的分子机制奠定了理论基础。
猜你喜欢
云南农业(2021年11期)2021-11-12
信阳农林学院学报(2021年1期)2021-04-01
林业科技(2020年3期)2021-01-21
今日农业(2020年13期)2020-12-15
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
现代农业科技(2017年13期)2017-08-09
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
农村农业农民·B版(2016年7期)2016-10-21
