
基于CLIMEX的2种核桃蚜虫潜在适生区分析
2020-07-07 07:16:26马德英刘孝贤高桂珍吕昭智新疆农业大学农学院新疆乌鲁木齐8005中国科学院新疆生态与地理研究所新疆乌鲁木齐800新疆农业大学林学与园艺学院新疆乌鲁木齐8005青岛农业大学植物医学学院山东青岛6609
生物安全学报 2020年2期
王 婷, 马德英, 刘孝贤, 张 苹, 高桂珍, 吕昭智,*新疆农业大学农学院,新疆乌鲁木齐8005;中国科学院新疆生态与地理研究所,新疆乌鲁木齐800;新疆农业大学林学与园艺学院,新疆乌鲁木齐8005;青岛农业大学植物医学学院,山东青岛6609
生物入侵是全球生物多样性丧失的主要原因之一,对各国生态系统、环境和社会经济构成的威胁日益严峻(蒋小龙等,2018)。外来入侵生物一旦成功建立种群后,根除难度非常大;预判和预测未来有害生物的入侵及其过程,对采取有效的防控措施,防止其进一步传入和扩散,避免造成更大的经济损失具有重要的参考价值。目前,生态位模型已成功应用到多个物种的管理和风险评估上,对入侵物种的管理和指导具有重要的参考价值(王聪等,2018)。
核桃Juglans regiaL.起源于中亚,世界各地都有大面积的种植(Martínezet al.,2010)。 据统计,2017年,全球核桃种植面积109.77万hm2,我国核桃种植面积占44.63%。核桃蚜虫包括核桃全斑蚜Panaphis juglandisGoeze与核桃黑斑蚜Chromaphis juglandicolaKaltenbach,2种蚜虫为单食性,是危害核桃的主要刺吸式害虫。核桃黑斑蚜在大部分核桃产区已有分布,2018年核桃全斑蚜在我国新疆伊犁和贵州陆续被发现,是我国新入侵的有害生物,具有潜在的危险性和扩散能力(吴跃开等,2018;邢海超等,2018)。
寄主核桃在世界范围内大面积分布,包括北美洲、欧洲大部分国家以及亚洲部分国家和地区,预测核桃蚜虫在全球范围内的潜在分布,对未来核桃有害生物的防治有重要价值。已有大量证据表明,气候在迅速的变化,政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)第四次评估报告(IPCC,2007)指出,1990—2100年之间,全球平均地表温度预计会升高2.4~6.4℃。本研究基于2种核桃蚜虫已有的研究数据,包括生活史、生物学特征和已知分布,选用以气候条件决定生物种群地理分布和数量变化的CLIMEX模型模拟2种核桃蚜虫在不同气候条件下的潜在分布,比较气候变化对2种核桃蚜虫分布的影响,以期为核桃种植和管理提供依据。
1 材料与方法
1.1 CLIMEX模型
CLIMEX是Suthrest和Maywald在1985年建立,通过物种在已知地理分布区域的气候参数来预测物种的潜在地理分布和相对丰度,并能分析气候对物种生长发育影响,能够比较全面地评估生物在新栖息地适生能力的一个动态模拟模型(宋红敏等,2004)。在CLIMEX模型中,有2个基本假设:(1)气候是影响物种分布和相对丰度的主要因素;(2)该物种在1年内经历适宜种群增长时期和不适合甚至危及生存这2个时期(贾文明等,2005)。
CLIMEX预测的物种分布由多个生物学参数非线性联合作用。通过调节参数可生成生态气候参数(ecoclimatic index, EI),由胁迫指数(stress index,SI)、生长指数(growth index,GI)和滞育、有效积温综合计算生成,表示物种的适生程度。区域EI值越大,表示物种在该地区越适合生存;胁迫指数(冷胁迫、热胁迫、干胁迫、湿胁迫)以及相互关系描述了物种在不良气候条件下的生存能力,该指数主要是限制种群的分布界限;GI类似于种群内禀增长率,表明了物种在理想条件下的增长速度和物候特点,GI值越大,表示物种在此条件下增殖潜力越快(Kriticoset al.,2014)。目前,CLIMEX模型已被用于估计当前和未来气候下多种入侵物种的潜在分布和种群模拟(孔令斌等,2008; Chejaraet al.,2010; Poutsmaet al.,2008; Sutherst&Bourne,2009)。
1.2 基础数据
1.2.1 寄主分布 核桃分布在欧洲、美洲和亚洲。在世界的50多个国家都有种植,主要种植国家有中国、美国、伊朗、土耳其、乌克兰、墨西哥、印度、巴基斯坦、智利、法国、罗马尼亚、希腊、埃及等(张毅萍,2001; Martínezet al.,2010)。 我国主要核桃产区包括新疆、青海、甘肃、宁夏、陕西、山西、天津、辽宁、山东、湖南、贵州、四川、广西等地(张毅萍,2001; Fenget al.,2018)。
1.2.2 核桃蚜虫的生物学参数 基于本课题组前期对蚜虫生命表的研究,得到参数 DV0、DV1、DV2、DV3、SM0、SM1、SM2、SM3,其他参数指标参考相关文献进行拟合确定(马菲等,2014;Avilaet al.,2019)。经过反复模拟和调试,模拟的分布覆盖了已有的蚜虫记录点,最终确定了CLIMEX模型的22个相关参数(表1)。
1.2.3 2种核桃蚜虫已知地理分布 核桃黑斑蚜原产于中东和中亚(Leeet al.,2018),国外分布于中亚、中东、非洲、丹麦、瑞典、西班牙、英国、德国、波兰与北美等地区(Aqaverdi&Inqilab,2018;Heie,1982; Leeet al.,2018)。 我国 1986 年在辽宁、山西、北京相继发现(杨俊杰和郭德明,1998)。
核桃全斑蚜原产于伊朗(Coeuret al.,2010),广泛分布在欧洲的西班牙、意大利、希腊、丹麦和瑞典等地(Aqaverdi&Inqilab,2018)。1928年首次出现在美国俄勒冈州威拉米特河谷;1952年又在加利福尼亚州圣西荷附近的核桃树上发现,次年,核桃全斑蚜蔓延到旧金山湾地区的大部分核桃产区以及美国的拜伦(Olson,1974; Paulsenet al.,2013);随后在欧洲、亚洲很多国家陆续被报道(吴跃开等,2018;邢海超等,2018; Heie,1982; Leeet al.,2018)
1.2.4 气候数据 选取的气候模型以1975年为中心的30年的气候数据,空间分辨率为10′,包括未来2030、2080年的气候情景,月平均降水量、日最高气温、日最低气温、9:00和15:00的空气相对湿度(Kriticoset al.,2014)。温室气体排放场景具体参考IPCC(2007)第四次评估报告中基于各种能源供应和利用技术发展速度相当的假定条件下,不过分依赖于某一特定能源资源的利用情景。

表1 核桃黑斑蚜与核桃全斑蚜CLIMEX参数值Table 1 CLIMEX parameter values of C.juglandicola and P.juglandis
1.3 研究方法
运用CLIMEX的"Compare locations"功能,分别得到核桃全斑蚜和核桃黑斑蚜现在与未来全球范围内潜在分布地区的EI值,将EI值划分为3个等级:1≤EI<5为低度适生区;5≤EI<15为中度适生;EI≥15为高度适生区。EI值越大则表示该地区气候环境越适合核桃蚜虫生存。利用美国环境系统研究公司(Environmental Systems Research Institute,ESRI)开发的ArcGIS 10.2软件的反距离加权插值分析功能得到全球核桃种植区分布及核桃蚜虫的潜在气候适生区。在模拟的基础上,与核桃树的分布、核桃蚜虫的已知分布进行验证,并与核桃蚜虫的实际种群动态进行对比,评估和提高计算模型的准确性,最终计算和模拟出2种核桃蚜虫适生区的差异性及其未来气候条件下适生区的变化。
2 结果与分析
2.1 目前气候条件下2种核桃蚜虫的气候适生区
研究表明,世界范围内2种核桃蚜虫适生区主要分布在温带及亚热带北部地区。国外的适生区分布在欧洲大部分地区,中亚、西亚、东亚的季风气候区,北美洲东部沿海地区和中部大平原地区,南美洲南部的少数地区。在我国主要集中分布在东北、华北和华中地区,另外在我国的西北地区和西南山区有零星分布。从CLIMEX模型模拟计算出的适生区来看,2种核桃蚜虫潜在分布区与世界核桃主要产区是相重叠的(图1)。
在我国,核桃黑斑蚜低度适生区在内蒙古东部及中部地区、黑龙江北部、河北北部、山西北部、陕西北部、甘肃南部部分区域、四川部分区域、新疆北部边疆地区;中度适生区在黑龙江、吉林中东部地区、辽宁西部、山西中部、陕西中部、宁夏南部、甘肃南部;高度适生区在辽宁中部及辽东半岛地区、山东的黄河以南地区、山西南部、陕西南部、甘肃陇东地区、江苏长江以北地区、湖北北部、重庆和四川的大巴山区、云南云贵高原北部地区。
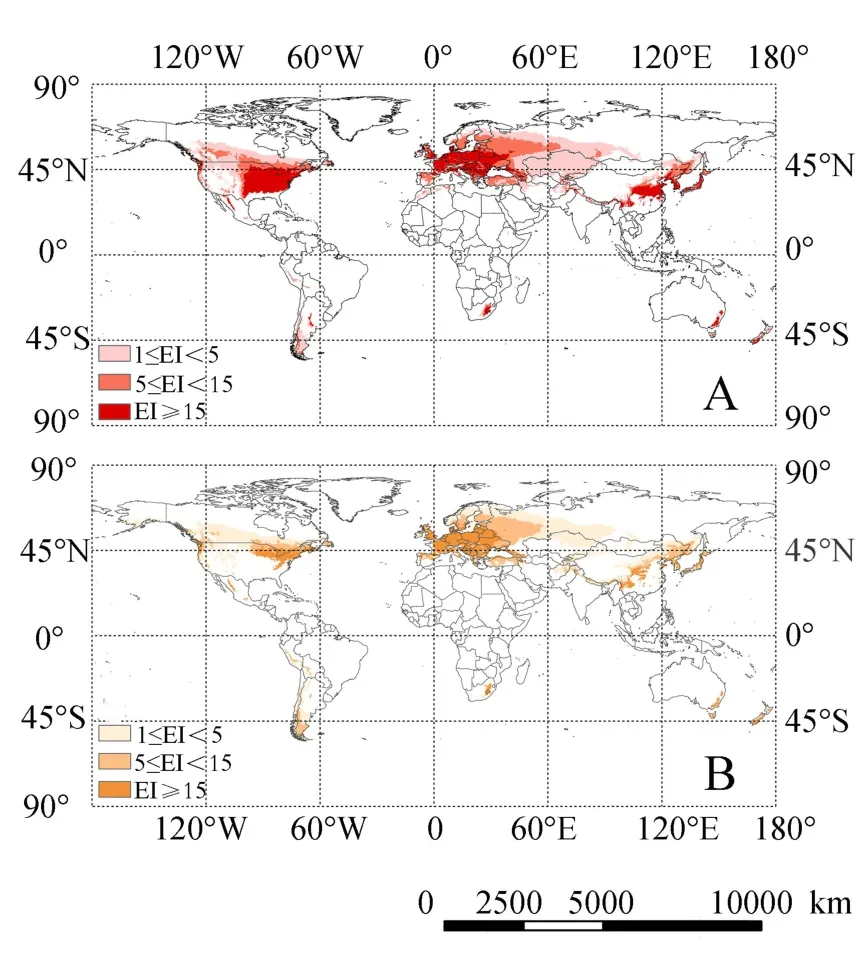
图1 目前核桃黑斑蚜(A)、核桃全斑蚜(B)的潜在适生区Fig.1 The current potential distribution of C.juglandicola(A)and P.juglandis(B)on walnut throughout world
世界范围内,核桃黑斑蚜低度适生区:在亚洲,分布于俄罗斯西伯利亚南部地区、哈萨克斯坦、乌兹别克斯坦北部、伊朗及阿富汗北部,土耳其;在欧洲,分布在俄罗斯东欧平原北部地区、瑞典、芬兰、英国北部、西班牙、乌克兰东部;在非洲,分布于阿尔及利亚、摩洛哥北部,南非也有少量分布;南美分布在秘鲁、智利及阿根廷南部少数区域。中度适生区:在亚洲,分布在塔吉克斯坦、乌兹别克斯坦、阿富汗;在欧洲,分布在瑞典、芬兰、英国北部、西班牙、乌克兰;在非洲,分布在阿尔及利亚、摩洛哥北部。高度适生区:在亚洲,分布在韩国、朝鲜、尼泊尔;在欧洲,高度适生区域面积比例较大,包括乌克兰西部、波兰、德国、意大利、法国、捷克、罗马尼亚、奥地利、丹麦、荷兰、英国南部、保加利亚、南斯拉夫、匈牙利等国家;北美洲主要潜在分布区在加拿大南部和美国。
与核桃黑斑蚜相比较,核桃全斑蚜为我国新入侵的害虫,低度气候适生区范围更窄,中、高度气候适生区范围则较广。核桃全斑蚜在我国内蒙古无适生区域,华北、东北地区中低度适生区也大幅度减少,高度适生区则与核桃黑斑蚜相似,在新疆西北部出现中度适生区;国外在欧洲及北美洲中、高度适生区则比核桃黑斑蚜广。
2.2 未来气候条件下2种核桃蚜虫在世界范围的气候适生区
由CLIMEX模型模拟计算出2030、2080年2种核桃蚜虫的气候适生区(图2)。核桃黑斑蚜2030年气候条件下世界范围内的适生区域范围相比目前有扩大趋势。亚洲适生区无明显变化,与目前气候条件下的适生区相似。欧洲气候适生区显著扩大,在挪威、瑞典、芬兰、俄罗斯的中、高度适生区均扩大。北美洲的低、中、高度适生区也有部分增加。在2080年气候条件下,核桃黑斑蚜的气候适生区域范围相比2030年表现为扩大趋势。欧洲、北美洲的气候适生区纬度明显升高,低、中、高度气候适生区都向北移动,亚洲的气候适生区与2030、2080年气候条件下相比,中、高度气候适生区也有部分增加。
在2030年气候条件下,核桃全斑蚜的气候适生区域范围相比目前气候条件下也表现为扩大趋势。在亚洲的气候适生区中,俄罗斯西部的低、中度适生区明显扩大,其他国家的气候适生区与目前气候条件下的气候适生区相似。欧洲、北美洲的气候适生区也明显扩大;2080年气候条件下,核桃全斑蚜的气候适生区范围相比2030年表现为扩大趋势。在亚洲,俄罗斯西部的中、高度气候适生区大幅度增加,其他国家的气候适生区与2030年气候条件下的气候适生区相似;在欧洲,气候适生区分布的纬度明显升高,并且多为高度适生区。北美洲的中、高气候适生区也有部分增加,加拿大的中、高度气候适生区增加,美国的高度气候适生区减少。
2.3 未来气候条件下核桃蚜虫在中国的气候适生区
模拟结果表明,我国核桃蚜虫的分布区在目前、2030、2080年情景下存在一定的差异性,随气候的变化核桃黑斑蚜未来潜在气候适生区范围西部增加、中部减小,东部增加;核桃全斑蚜未来潜在气候适生区范围为西部增加、中部减小,东部无明显变化。
核桃黑斑蚜2030年气候条件下的气候适生区与目前相比,东北、华北地区适生区面积均增大,高度适生区面积在黑龙江佳木斯,吉林长春,辽宁沈阳、本溪、丹东明显增加,新疆西北部的中、低度适生区面积也表现为增大。2080年气候条件下的适生区域范围相比2030年,东北地区的中、高度适生区面积均增大;江苏、安徽、河南的高度适生区也减小,表现为华北、华中、华东地区的适生区面积变窄;新疆西北部的中、低度适生区面积表现为扩大。
核桃全斑蚜2030年气候条件下的气候适生区与目前相比,在华北、华中,安徽、江苏、河南的适生区面积减少,新疆西北部的中度适生区面积增大。2080年气候条件下气候适生区范围与2030年相比,东北地区中、高度适生区面积减小;江苏、安徽、河南的高度适生区也减小,表现为华北、华中、华东地区的适生区面积变窄并小于同期核桃黑斑蚜适生区。
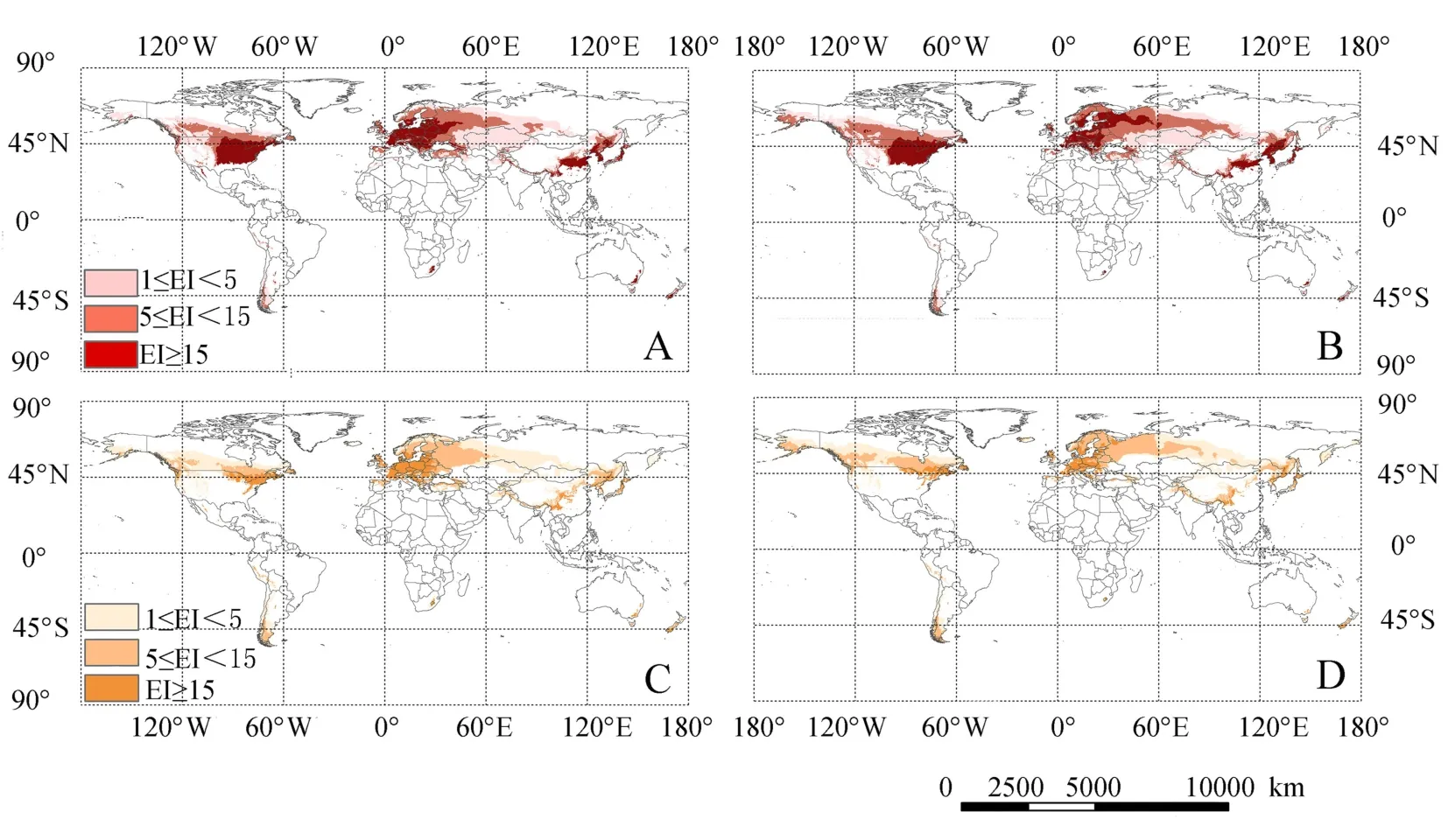
图2 未来条件下核桃黑斑蚜2030(A)、2080年(B),核桃全斑蚜2030(C)、2080年(D)的潜在气候适生区Fig.2 The potential distribution of C.juglandicola in 2030(A),2080(B)and P.juglandis in 2030(C),2080(D)
2.4 目前气候下2种核桃蚜虫种群动态分析
从目前气候下CLIMEX模型模拟的我国部分地区的GI值来看,核桃黑斑蚜在辽宁、山东、河南、新疆的GI值均呈双峰型,说明核桃黑斑蚜在这些地区一年内有2个高峰期。而核桃全斑蚜在部分地区GI值为单峰型,在部分地区则为双峰型。2种蚜虫的种群动态与当地气候条件(特别是夏季的高温)和蚜虫生物学特征(耐高温的能力)密切相关(图 3)。
3 结论与讨论
在CLIMEX模型预测中,在目前气候条件下,核桃黑斑蚜的中、高度潜在适生区面积明显大于核桃全斑蚜。在未来气候条件下,2种核桃蚜虫的潜在气候适生区纬度越来越高,这可能是全球气候变暖导致的结果。大多数植物和动物(变温动物)的分布都是由气候决定的(Andrewartha&Birch,1954; Woodward,1987),Eagar(1978)观察到凤仙花Impatiens balsaminaL.在高温下表皮干燥致使凤仙花蚜虫Adelges piceaeRatz.死亡,石作雄(2019)在玉米田中调查发现夏季多雨,湿度过大,会导致蚜虫大量死亡。本研究结合对2种核桃蚜虫的基础生物学分析发现,核桃黑斑蚜更倾向于温暖的环境,核桃全斑蚜更倾向于湿润的环境,这可能是影响2种核桃蚜虫的潜在分布的环境因子。
CLIMEX模型中每个参数值的更改都会导致模拟结果的不同,在调试的22个相关参数中,影响核桃蚜虫地理分布的的主要胁迫因子是干胁迫;从2种核桃蚜虫在气候带分布来看,卵滞育是其分布的关键因子之一,昆虫的滞育主要发生在高纬度寒冷的冬季或热带、亚热带地区干旱、炎热的季节(曾庆礼等,2004)。2种核桃蚜虫主要分布在温带,在冬季以卵滞育越冬。在未来气候向着温暖、湿润的方向发展条件下(秦大河等,2005),其越冬习性可能发生改变,分布区也可能延伸到热带等目前无适生性的区域,这需要更一步的研究和分析。

图3 目前气候条件下核桃黑斑蚜、核桃全斑蚜在中国部分适生区的生长指数Fig.3 GI values of C.juglandicola and P.juglandis in the suitable areas of China under current climatic conditions
2种核桃蚜虫年际内的种群动态存在很大的差异性,特别是种群波动存在单峰或者双峰现象,而影响这种现象出现的原因尚不清楚。1年内核桃黑斑蚜种群有2个危害高峰,一般出现在6月和8月中下旬到9月初左右(杨俊杰和郭德明,1998;Jaskiewicz&Kmeic,2007)。昆虫种群动态可能受到密度依赖性、复杂营养相互作用(Rosenheim,1998)、气候和植物条件、迁移和景观(Deet al.,1991;Kareiva,1990)等因素的综合影响。研究发现,在加利福尼亚的中央谷,6月份核桃黑斑蚜较多(>5蚜虫·叶-1)的核桃果园,9月份基本没有蚜虫;6月份蚜虫数量较少的地块(<5蚜虫·叶-1)并没有表现出如此一致的趋势,有些地块在9月份蚜虫密度较高(Mace&Mills,2017),这与 Sluss(1967)观察到核桃黑斑蚜在叶片上的密度受先前蚜虫取食叶片使其发生化学变化的影响相一致。导致核桃黑斑蚜种群出现双峰的原因还可能与夏季核桃的休眠引起植物营养下降以及这一段时期的高温相关。但全斑蚜种群动态相对比较复杂,Jaskiewicz&Kmeic(2007)在波兰发现核桃全斑蚜种群出现单峰,Atlihanet al.(2015)研究表明,核桃全斑蚜种群动态在土耳其呈双峰模式,但其内在因子需要严格的试验证实。
人类活动是核桃蚜虫传播的重要因素。在地中海沿岸的欧洲地区,中亚和中国东北沿黄河流域这3个早期主要果树栽培中心的宏观区域中,发现了核桃种子的化石(Wilkinsonet al.,2012; Xi,1989),而核桃黑蚜虫原产于中东和中亚,核桃全斑蚜原产于伊朗,这与早期核桃种植区相吻合。随后,丝绸之路等贸易往来将中国和印度连接到地中海地区,促进了核桃蚜虫的传播扩散,Pollegioniet al.(2015)研究发现,人类活动在较大地理区域上的相似性促进了核桃的传播,核桃蚜虫随之从早期的地理分布区向其他区域扩散。
目前,国外核桃蚜虫发生区多使用化学农药进行防治,引起农药抗性倍数急剧上升,天敌多样性下降(Van denet al.,1979)。单独依靠农药很难控制蚜虫种群增长和成灾(Dixon&Kindlamnn,1998)。而在我国,核桃蚜虫在部分核桃种植区发生,经济损失相对较少,不建议进行化学防治,特别是利用飞机进行大面积的化学防治。如在新疆地区,核桃分布区约是棉花分布区的1/6,棉田中的自然天敌,如异色瓢虫Harmonia axyridisPallas、大草蛉Chrysopa pallens(Rambur)、食蚜蝇 Syrphidae等天敌迁移到果园可对核桃蚜虫进行控制;在云南地区,丰富的作物资源,多样性的天敌群落,使核桃蚜虫可在自然条件下得到控制。
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
小天使·一年级语数英综合(2022年5期)2022-05-25 16:36:27
当代水产(2022年2期)2022-04-26 14:25:48
发明与创新·小学生(2021年5期)2021-08-23 02:24:00
发明与创新(2021年19期)2021-05-07 07:53:00
当代水产(2020年4期)2020-06-16 03:22:52
当代水产(2020年4期)2020-06-16 03:22:46
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
小学生导刊(2018年31期)2018-12-06 08:36:50
