
饲料脂肪水平与黄颡鱼幼鱼生长、体营养成分及肌肉脂肪酸组成的关系
2020-07-04 00:57冯鹏霏何金钊马华威潘传燕孔丽芳
饲料工业 2020年12期
冯鹏霏 何金钊 马华威 潘传燕 滕 云 孔丽芳 陈 涛
(1.广西壮族自治区水产科学研究院,广西南宁530021;2.广西壮族自治区水产引育种中心,广西南宁530031;3.南宁学院,广西南宁530299)
脂肪是鱼类饲料中的重要成分,为鱼体提供能量和必需脂肪酸,鱼类的正常生长发育离不开脂肪营养。脂肪可以节约蛋白质及减少氮排泄[1-2],鱼体生长离不开脂肪,但如果饲料中脂肪含量过高,则也会影响鱼体正常的生长繁殖,导致鱼体脂肪沉积增加而发生疾病[3],所以不同品种的鱼类,其饲料应提供适宜含量的脂肪。
饲料中脂肪或脂肪酸对黄颡鱼生长[4-5]、体色[6]、酶基因[7]、肝脏脂肪积累[8]、血糖水平和酶活力[9]、卵巢脂类代谢[10]、生理因子[11]、生理代谢[12]等方面的影响研究已经有很多报道,亦有黄颡鱼脂肪需求量[13-14]研究,但对饲料中不同脂肪水平对黄颡鱼幼鱼生长、体营养成分及肌肉脂肪酸组成影响的研究鲜有报道,本报道以黄颡鱼幼鱼为研究对象,在饲料中添加不同水平的豆油,分析饲料中不同脂肪水平对黄颡鱼幼鱼生长、体营养成分及肌肉脂肪酸组成的影响,以确定黄颡鱼幼鱼饲料中适宜的脂肪添加水平,为黄颡鱼人工饲料配方的进一步优化提供数据参考。
1 材料与方法
1.1 试验材料
试验用黄颡鱼幼鱼由南宁市朗多畜禽水产养殖专业合作社提供,幼鱼平均体质量(1.43±0.09) g。
试验鱼暂养6 d,饥饿24 h,选择的健康幼鱼(体质量大致相同)900尾,分为5组,每组设3个平行,每个平行60尾,在室内水泥池(1 m×1 m×0.8 m,每个水泥池为一个平行)中饲养60 d。试验鱼每天8:00和18:00分别饲喂试验饲料1次,试验1~30 d每天投喂饲料量为鱼体重的5%,31~60 d投喂饲料量以试验鱼15 min内吃完为准。试验期间平均水温29~32 ℃,DO(7.23±0.44) mg∕l,pH值(7.1±0.6),NH3-N(0.04±0.01) mg∕l。
试验饲料以鱼粉、玉米粉等为基础料,以豆油调节脂肪水平,配制成5 组饲料,各组的豆油添加量分别为0%(1 组)、2%(2 组)、3.5%(3 组)、5%(4 组)和6.5%(5 组)。饲料组成及营养水平见表1,各组饲料的脂肪酸组成见表2。
饲料制作:先把基础料、添加剂和油脂混合好,然后加水搅拌均匀,揉成团状后逐个投入饲料制粒机,制成条状饲料(直径约1.5 mm),放入60 ℃鼓风干燥箱中持续烘干6 h,然后制成长2 mm 左右的颗粒饲料,密封冷藏保存。
1.2 样品采集与分析方法
1.2.1 生长指标和鱼体常规营养成分的分析测定
试验结束后,鱼体肌肉取样,分别采用烘干法、索氏抽提法、凯氏定氮仪及马弗炉高温灼烧法测定幼鱼肌肉中的水分、粗脂肪、粗蛋白及粗灰分含量。计算幼鱼成活率、饲料系数及增重率。
SR(%)=100×终末尾数∕初始尾数
FCR=F∕(Wt-W0)
WGR(%)=100×(Wt-W0)∕W0
式中:W0——初均重(g);
Wt——末均重(g);
WGR——增重率(%);
FCR——饲料系数;
F——摄食饲料总量(g);
SR——成活率(%)。
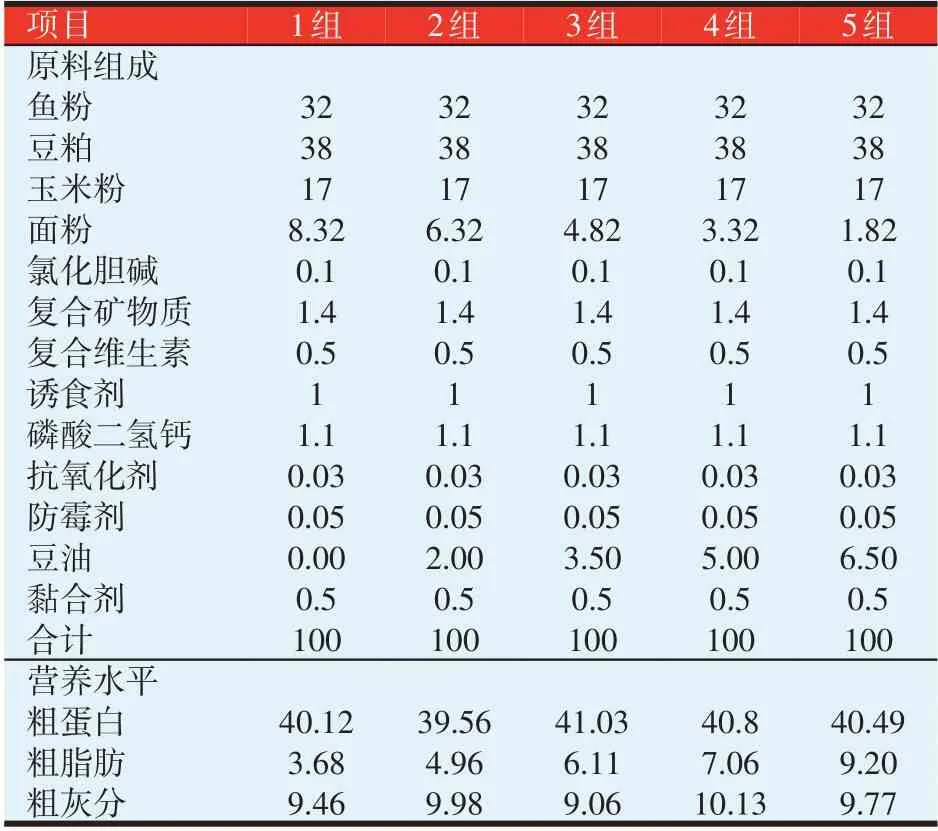
表1 试验饲料组成及营养水平(%)
1.2.2 脂肪酸组成的测定
取鱼体背部肌肉1 g 左右,粉碎后置于研钵中研磨,研钵中加5 ml Folch液并研磨肌肉,将研磨至糜状的肌肉倒入500 ml 的圆底烧瓶,加15 ml Folch 液于烧瓶中,置于55 ℃水浴锅中回流2.5 h,滤去残渣后,将烧瓶中的澄清液体用25 ml 容量瓶定容,取其中10 ml 液体放入80 ml 小圆瓶中,99.9%氮气吹干小圆瓶中的液体,然后加入2 ml 0.5 mol∕l的氢氧化钾甲醇溶液,充氮气99.9%,置于50 ℃水浴锅中振摇3 min左右,待小圆瓶内的油珠消失后,用自来水冲淋冷却瓶身;继续往小圆瓶内加2 ml 10%的三氟化硼乙醚溶液,轻轻摇匀并充氮气,置于50 ℃水浴锅中静置约3 min,以自来水冷却,加2 ml正庚烷,充分振摇摇匀,在常温下静置约5 min,用饱和食盐水溶液淋洗上清液,静置待液体分层,取少量上清液置于1 ml EP 管中,加少量无水硫酸钠防潮。取微量上清液进气相色谱仪,测定肌肉样品中脂肪酸种类及相对含量。
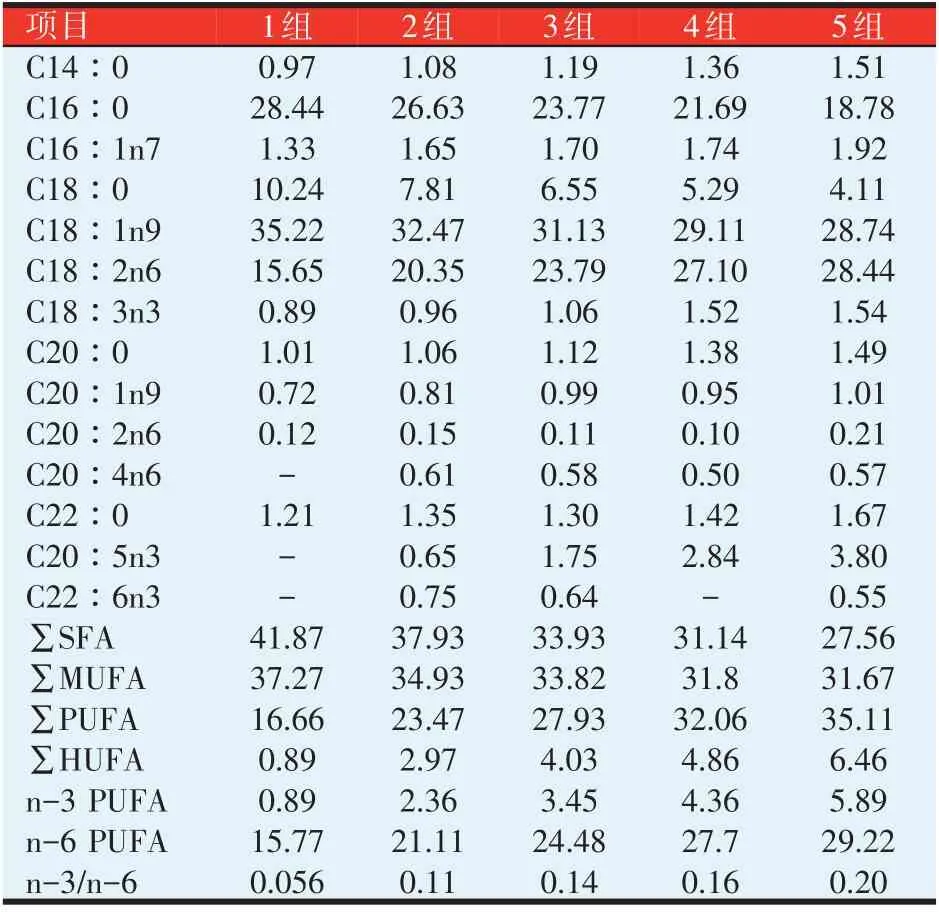
表2 各组饲料的脂肪酸组成(%)
采用气相色谱法测定鱼体脂肪酸组成。使用美国Sigma 公司生产的脂肪酸甲酯标准品。澳大利亚SGE公司的规格为60 m×0.25 mm的BPX70石英毛细管柱,220~240 ℃程序升温;汽化室和FID 温度为290 ℃;流速60 ml∕min 的氮气作为载气,60 ml∕min H2,600 ml∕min空气,50 ml∕min尾吹气;氢离子火焰检测器;进样量1 μl,分流比100∶1。按照峰面积归一法计算各脂肪相对含量。
1.3 数据处理
试验结果整理后统一录入Excel 表格,统计软件SPSS 19.0 对数据进行单因子方差分析,试验结果以“平均数±标准差”表示,采用Duncan's 多重比较法分析平均数的差异显著性(P<0.05)。
2 结果与分析
2.1 不同脂肪水平对黄颡鱼幼鱼生长指标的影响(见表3)
表3中各组间试验鱼初均重、存活率均差异不显著(P>0.05)。黄颡鱼幼鱼末均重及增重率以4 组最高,5组次之,1组最低,4、5组末均重显著高于其他各组(P<0.05),3 组显著高于1、2 组(P<0.05);增重率3、4、5 组显著高于1、2 组(P<0.05);饲料系数1 组最高(1.38),5 组次之,4 组最低,1、5 组显著高于2、3、4 组(P<0.05),2、3、4组之间无显著性差异(P>0.05)。

表3 不同脂肪水平对黄颡鱼幼鱼生长指标的影响
2.2 不同脂肪水平对黄颡鱼鱼体营养成分的影响(见表4)
表4 中各组鱼体肌肉水分、蛋白、灰分含量均无显著性差异(P>0.05)。其中2 组水分含量最高,3 组最低;肌肉蛋白含量以5 组最高,4 组最低;肌肉灰分含量3 组最高,5 组最低;肌肉脂肪含量以5 组最高(12.11%),1 组最低(10.19%),3、5 组显著高于2、4 组(P<0.05),2、4 组显著高于1 组(P<0.05),2、4 组之间,3、5组之间无显著性差异(P>0.05)。
2.3 不同脂肪水平对黄颡鱼肌肉脂肪酸组成的影响(见表5)
2.3.1 黄颡鱼幼鱼肌肉、饲料中各脂肪酸种类及相对含量的相关性
由表5 可知,黄颡鱼幼鱼肌肉组织中共检测出15 种脂肪酸。大类脂肪酸的相对含量高低顺序均为∑SFA>∑MUFA>∑PUFA>∑HUFA。主要的SFA 是C16∶0、C18∶0,主要的MUFA 则是C18∶1n7、C18∶1n9及C16∶1n7,主要的PUFA为C18∶2n6、C18∶3n3及C22∶6n3,主要的HUFA是C18∶3n3及C22∶6n3。
饲料脂肪酸组成各组和试验鱼肌肉脂肪酸组成有很大相关性。随着脂肪添加量的增加,各组饲料中的C16∶0、C18∶0、C18∶1n9、SFA以及MUFA相对含量迅速降低,C18∶2n6、C20∶5n3 以及PUFA、HUFA相对含量则迅速提升;各组试验鱼肌肉C18∶2n6、MUFA、PUFA和HUFA亦呈现出相同的变化趋势。各组饲料及试验鱼肌肉组织中的n-3∕n-6均低于0.33。
2.3.2 黄颡鱼幼鱼肌肉组织和饲料中脂肪酸组成的差异性
4、5 组饲料中大类脂肪酸相对含量大小顺序为∑PUFA>∑MUFA>∑SFA,4、5 组试验鱼肌肉中的大类脂肪酸相对含量大小顺序则为∑SFA>∑MUFA>∑PUFA,肌肉组织大类脂肪酸组成跟饲料有所不同;随着脂肪添加量的增加,各组饲料中的C16∶0、C18∶0、C18∶1n9、SFA和MUFA相对含量迅速降低,C18∶2n6、C20∶5n3、PUFA和HUFA相对含量迅速提升,而试验鱼肌肉C16∶0、C18∶0、C18∶1n9、C20∶5n3、SFA相对含量变化不大,表明鱼体组织脂肪酸相对含量不一定随着饲料变化而变化。
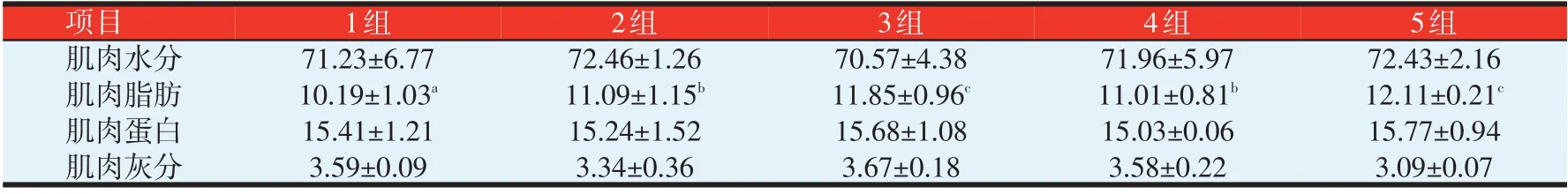
表4 不同脂肪水平对黄颡鱼幼鱼鱼体营养成分的影响(%)
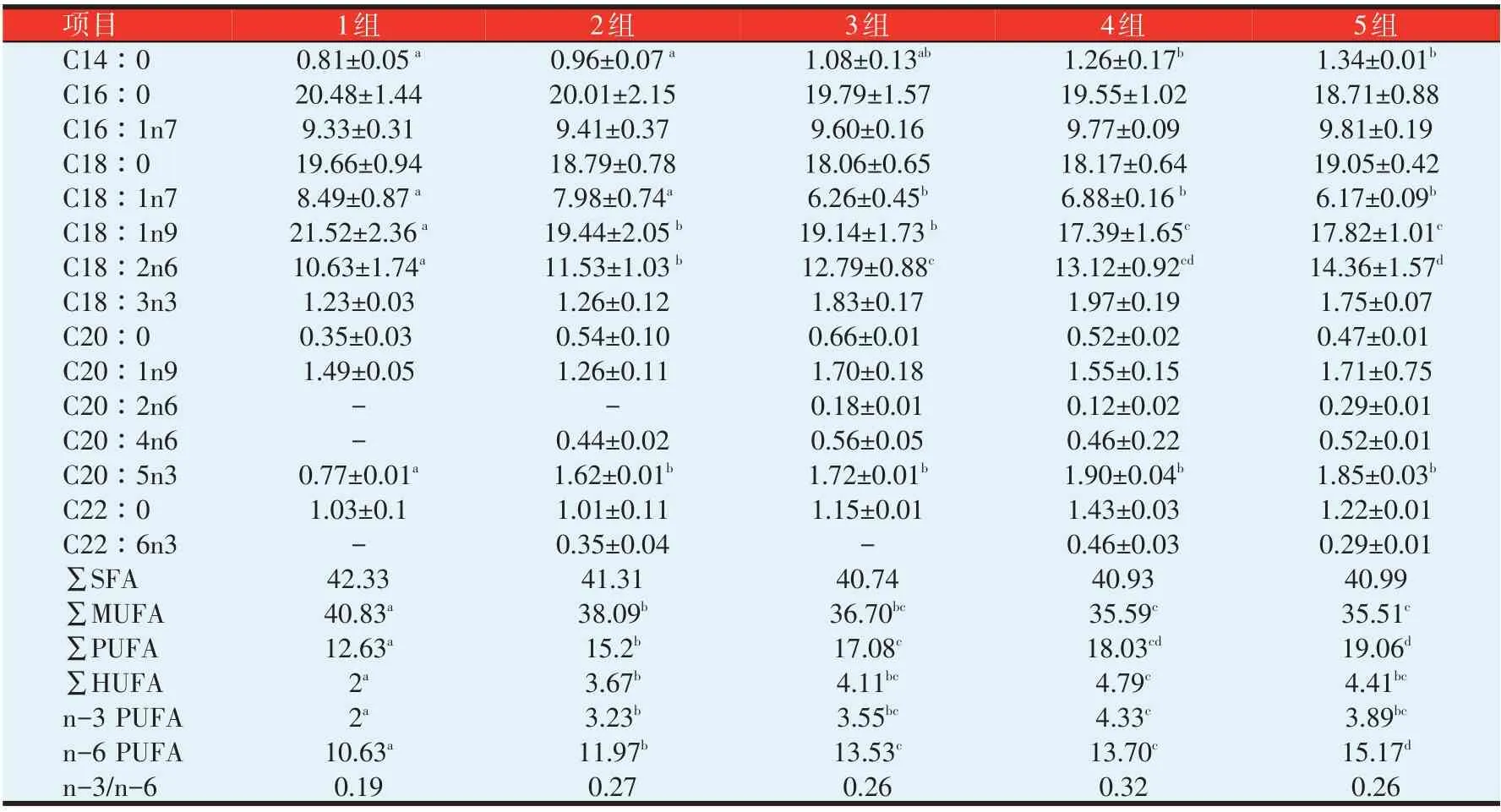
表5 不同脂肪水平对黄颡鱼幼鱼肌肉脂肪酸组成的影响(%)
黄颡鱼幼鱼肌肉中大类脂肪酸与饲料中相比,肌肉组织中的SFA 及MUFA 相对含量均高于相应饲料。黄颡鱼幼鱼肌肉中C18∶1n9相对含量远低于饲料,C16∶1n7 相对含量则远高于饲料,肌肉中C18∶1n7 相对含量达8%以上,饲料中没有检出C18∶1n7。1~5组黄颡鱼幼鱼肌肉中的PUFA 及n-6 PUFA相对含量均低于相应饲料;1~3组黄颡鱼幼鱼肌肉中的n-3 PUFA 相对含量高于相应饲料,4~5 组则低于相应饲料。鱼体肌肉C18∶2n6相对含量均比相应饲料中低的多,C18∶3n3 相对含量则是鱼体肌肉稍高一些。3、4、5 组饲料中C20∶5n3(EPA)的相对含量均比相应鱼体高,C22∶6n3 相对含量肌肉和饲料中相差不大。各组肌肉n-3∕n-6高于相应饲料。
2.3.3 黄颡鱼幼鱼肌肉组织中各类脂肪酸的相对含量
比较各组黄颡鱼肌肉的脂肪酸相对含量,发现5组的C14∶0含量最高(1.34%),4、5组显著高于1、2组(P<0.05),1、2、3组之间差异不显著(P>0.05);各组试验鱼之间肌肉C16∶0、C16∶1n7、C18∶0、C18∶3n3相对含量差异不显著(P>0.05),4组试验鱼肌肉C18∶3n3相对含量最高(1.97%),1组最低(1.23%);3、4、5组试验鱼肌肉C18∶1n7相对含量显著高于1、2组,1、2组之间,3、4、5组之间无显著性差异(P>0.05);4、5 组试验鱼肌肉C18∶1n9相对含量显著高于2、3组(P<0.05),2、3组显著高于1组(P<0.05);C18∶2n6以5组最高(14.36%),5组显著高于3组,3、4、5组显著高于1、2组(P<0.05),2 组显著高于1组(P<0.05);除1组外,各组试验鱼肌肉C20∶4n6相对含量比较接近;1组C20∶5n3(0.77%)相对含量显著低于其他各组(P<0.05),其他各组间差异不显著(P>0.05),4组最高(1.90%);C22∶6n3相对含量以4组最高(0.46%),其他各组含量较低或未检出。
比较各组鱼肌肉中大类脂肪酸,∑SFA、∑MUFA均以1组最高,1组∑MUFA显著高于其他各组(P<0.05);∑PUFA以5组最高(19.06%),3、4、5组显著高于1、2组;∑HUFA、∑n-3 PUFA及∑n-6 PUFA在各组间存在不同差异,除∑n-6 PUFA外,均为4组最高,1组最低;n-3∕n-6以1组最低(0.19%),各组之间差异不显著(P>0.05)。
3 讨论
3.1 不同脂肪水平对黄颡鱼幼鱼生长指标的影响
饲料中脂肪含量过低,供给鱼体的能量就会减少,只能消耗鱼体本身的蛋白质以维持能量供应,导致鱼体生长性能下降;若脂肪过量,则鱼类肝胰脏等组织会蓄积多余的脂肪在其中,诱导鱼体发生脂肪肝病变,最终影响鱼类正常生长[15]。
张丽燕等[13]以不同脂肪水平的饲料饲养平均体质量10.32 g的黄颡鱼100 d,发现饲料脂肪水平对黄颡鱼增重、特定生长率均无显著影响,但显著改变了其饲料系数;本试验中黄颡鱼幼鱼(平均体质量约1.43 g)末均重、增重率及饲料系数均随饲料脂肪水平的变化而变化显著。
于丹等[4]以不同脂肪水平的饲料饲喂黄颡鱼幼鱼,发现随着饲料脂肪水平的提高,黄颡鱼幼鱼增重率先升后降,对黄颡鱼幼鱼的生长起促进作用的饲料脂肪添加量范围为7.59%~7.65%;韩庆等[14]以鱼油∕豆油(1∶1)为脂肪源,以脂肪添加量分别为5.21%、7.23%、9.27%、11.31%、13.33%、15.37%的6 组半纯化饲料饲喂的黄颡鱼幼鱼(体重约8 g)40 d,发现当饲料脂肪含量低于11.31%时,随着饲料脂肪水平的提高,鱼体相对增重率及饲料转化率逐渐增加,当脂肪水平高于11.31%时,随着饲料脂肪水平的增加,鱼体相对增重率及饲料转化率又逐渐降低,确定黄颡鱼饲料最适脂肪添加量为11.31%。本试验中,随着饲料脂肪水平的升高,各组试验鱼的末均重和增重率均呈现先增后降的趋势,其中4组(脂肪添加量5%)末均重及增重率均达最高(17.30 g和1 118.3%),1组最低,增重率和末均重都是3、4、5组显著高于1、2组(P<0.05),造成这种试验鱼生长速度先增后降现象的原因可能是过高的脂肪含量降低了鱼体的生长性能,这跟韩庆等的研究结果一致,但不同之处在于韩庆等得出的最适脂肪水平为11.31%,本试验得出最适脂肪添加量为5%,这可能跟黄颡鱼幼鱼不同的生长阶段有关。袁立强等[5]研究发现,随着饲料脂肪水平的提高,瓦氏黄颡鱼的特定生长率先增后降,与本试验结果一致。本试验饲料系数则是4组(1.19)最低,1组最高(1.38),变化趋势不明显。综上分析,4组鱼体生长性能最好,3、5组次之,1、2组较差。
3.2 不同脂肪水平对黄颡鱼幼鱼肌肉常规营养成分的影响
高脂饲料促进了脂肪在鱼体肌肉中的积累。张世亮等[9]研究表明,随着糖∕脂肪比例的增加,全鱼粗脂肪含量会显著减少。郑珂珂等[7]研究表明,黄颡鱼肌肉营养成分随着饲料脂肪水平的改变而发生显著变化,其中肌肉脂肪含量随脂肪水平的增加而显著增加,肌肉粗蛋白含量则先升后降,饲料脂肪水平为7.9%和10.9%时能显著提高鱼体蛋白含量。本试验表明,除肌肉脂肪含量外,不同脂肪水平对黄颡鱼肌肉蛋白质、水分、灰分等常规营养成分均无显著影响影响。黄颡鱼幼鱼肌肉粗脂肪含量以5 组最高(12.11%),3 组次之(11.85%),1组最低(10.19%),3、5组显著高于其他各组(P<0.05),1组显著低于其他各组(P<0.05);与郑珂珂等[7]的研究结果不一样,本试验中黄颡鱼幼鱼肌肉蛋白及灰分含量呈现不规律的变化,肌肉蛋白质含量5组最高,水分含量2组最高,灰分含量3组最高。
3.3 不同脂肪水平对黄颡鱼幼鱼肌肉脂肪酸组成的影响
3.3.1 各组饲料脂肪酸组成和黄颡鱼幼鱼肌肉脂肪酸组成之间的相关性和差异性
鱼体摄取必需脂肪酸的一个重要途径是从饲料脂肪中获取,饲料脂肪酸组成一定会影响鱼体脂肪酸组成,鱼体脂肪酸的组成基本上反映了饲料脂肪酸组成特点[16-18]。本试验中,各组饲料中的C16∶0、C18∶0、C18∶1n9、SFA和MUFA相对含量迅速降低,各组试验鱼肌肉的MUFA相对含量也快速降低;各组饲料中C18∶2n6、C20∶5n3、PUFA和HUFA相对含量则迅速提升,各组试验鱼肌肉C18∶2n6、PUFA和HUFA亦迅速提升。各组饲料及试验鱼肌肉组织中的n-3∕n-6均低于0.33。
鱼体脂肪酸组成反映了饲料脂肪酸组成,但本试验中亦有很多种类脂肪酸的相对含量不一定随着饲料脂肪酸组成变化而变化,如各组试验鱼肌肉中C16∶0、C18∶0、C18∶1n9、C20∶5n3、SFA相对含量变化不大,未如饲料中相应的脂肪酸那样随着饲料脂肪含量的变化而迅速变化。再如4、5组试验鱼肌肉中的大类脂肪酸相对含量大小顺序则为∑SFA>∑MUFA>∑PUFA,不同于4、5组饲料中的顺序∑PUFA>∑MUFA>∑SFA。黄颡鱼幼鱼肌肉中C18∶1n9相对含量远低于饲料,肌肉中C16∶1n7相对含量则远高于饲料,肌肉中C18∶1n7相对含量达8%以上,饲料中没有检出C18∶1n7。肌肉C18∶2n6相对含量均比相应饲料中低的多。各组肌肉n-3∕n-6高于相应饲料。
本试验中黄颡鱼幼鱼肌肉中大类脂肪酸与饲料中相比,肌肉组织中的SFA及MUFA相对含量均高于相应饲料。这与刘飞[19]的结果不完全一致,刘飞研究不同脂肪源饲料对黄颡鱼脂肪酸组成的影响,结果表明,黄颡鱼肌肉SFA 相对含量低于饲料,而MUFA 相对含量高于饲料,认为黄颡鱼能将SFA去饱和后转化为MUFA,可能本试验中并不存在这样的转化。
C18∶2n6和C18∶3n3分别是n-6和n-3系列脂肪酸的前体,经碳链延长及去饱和作用后,C18∶2n6在机体内转化为HUFA[20]。本试验中,各组黄颡鱼幼鱼肌肉PUFA特别是n-6 PUFA的相对含量低于相应饲料,而C18∶2n6则远低于相应饲料,1、2、3组黄颡鱼幼鱼肌肉HUFA的相对含量又比相应饲料高,4、5组黄颡鱼幼鱼肌肉HUFA则比相应饲料低,可能在1、2、3组试验鱼体内也发生了类似转化,4、5 组未发生类似变化,至于C18∶2n6是在向DHA还是EPA转化,有待进一步研究。
陆游等[21]研究不同脂肪源饲料对黄颡鱼幼鱼生长性能、肌肉组织脂肪酸组成、血清生化指标及抗氧化能力的影响,发现各试验组黄颡鱼肌肉的C14∶0相对含量均低于相应饲料,而C16∶0、C18∶1n-9相对含量则高于相应饲料,认为黄颡鱼可能将多余的SFA通过去饱和或者延长碳链的方式生成C16∶0及C18∶1n-9,用于提供机体能量。而本试验中未出现这样的转化,黄颡鱼幼鱼肌肉中C14∶0 低于相应饲料,而C16∶0和C18∶1n9 相对含量亦低于饲料,C16∶1n7 相对含量则远高于饲料,饲料中没有检出C18∶1n7,但肌肉中C18∶1n7相对含量达到8%以上,可能C18∶1n9在黄颡鱼体内向C16∶1n7和C18∶1n7转化。
3.3.2 各组黄颡鱼幼鱼肌肉中脂肪酸组成的比较
鱼体正常生长所必需的脂肪酸可能是n-6 PUFA,富含HUFA的饲料亦能促进鱼体生长[22]。综合鱼体生长指标,4、5组黄颡鱼幼鱼生长性能较好,其增重率在1 031%~1 118%之间,饲料系数在1.19~1.36 之间,4 组增重率显著高于其他各组(P<0.05)。这可能跟4、5组鱼体肌肉中含有较高的C18∶2n6、C18∶3n3及EPA有关。
从营养学角度评价,亚油酸可能是有害的脂肪酸,Turchini等[23]研究发现,用植物油替代鱼油可能会导致亚油酸在鱼肌肉中大量沉积而难以移除。赵帅兵等[24]研究表明,使用棕榈油替代鱼油能降低亚油酸在黄颡鱼肌肉中的相对含量, 并且随棕榈油含量的增加,鱼体肌肉的n-3∕n-6比值显著提升,说明棕榈油替代鱼油可以改善及优化黄颡鱼的肌肉脂肪酸组成。本试验使用不同水平的豆油饲喂黄颡鱼幼鱼,其鱼体n-3∕n-6比值先逐渐增加,由0.19增加到0.32,随后降低至0.26,而肌肉亚油酸相对含量增幅不大,从10.63%到14.36%,说明饲料中添加一定水平的脂肪(豆油)能改善黄颡鱼幼鱼的肌肉脂肪酸组成,但过高水平的脂肪含量改善脂肪酸组成的作用有所下降。
猜你喜欢
化工管理(2022年13期)2022-12-02
浙江大学学报(理学版)(2022年1期)2022-02-21
农业工程学报(2021年4期)2021-05-09
中国生殖健康(2020年5期)2021-01-18
健康博览(2020年11期)2020-11-23
农业工程学报(2020年6期)2020-05-19
恋爱婚姻家庭·养生版(2018年11期)2018-11-26
养生保健指南(2018年4期)2018-04-11
养生保健指南(2016年12期)2017-01-06
食品工业科技(2014年13期)2014-03-11
