
137Cs-γ射线辐射对威灵仙种子发芽及幼苗生长的影响
2020-07-03 07:59:56刘志高梁立军季梦成
核农学报 2020年8期
叶 盈 刘志高 梁立军 季梦成
(浙江农林大学风景园林与建筑学院,浙江 杭州 311300)
威灵仙 (Clematis chinensis) 为毛茛科(Ranunculaceae)铁线莲属(Clematis.L)半常绿木质藤本植物,广泛分布于我国浙江、江苏、安徽、四川和云南等省[1]。 其花序圆锥状,腋生或顶生,萼片白色,花期6-9月,属多花、繁花类型,可用于家庭园艺栽培和园林垂直绿化,是极具开发潜力的本土铁线莲属观赏植物。
辐射育种是植物新品种培育的重要方法,被广泛应用于观赏植物育种和生长,并取得了一定成果。 在育种方面,蝴蝶兰花粉经80 Gy 剂量60Co-γ 射线辐射产生了一个黄花红唇红边的变异株系[2];宋恒等[3]用12 Gy 和16 Gy 剂量的60Co-γ 射线辐射杜鹃栽培种试管苗,获得了2 个花色突变系;刘春贵等[4]发现低于200 Gy 剂量的60Co-γ 射线辐射对路易斯安那鸢尾花色分离具有促进作用。 在生长方面,60Co-γ 射线对洋紫荆幼苗的苗高、地径和叶片分裂角度有明显的抑制作用[5]。 试验材料的成活率和变异性状出现的概率直接受辐射剂量的影响,较高剂量的γ 射线具有抑制作用,而低剂量的γ 射线可以促进细胞增殖、萌发、细胞生长和促进酶活性等[6-8],可见辐射对植物茎、叶及花的变异均有显著诱导作用[9]。 依据《植物新品种特异性、一致性和稳定性测试指南 铁线莲属》[10],叶、株型或花的形态变化均是新品种认定的重要指标。 观赏铁线莲品种常通过杂交和多代选育获得,仅有Nachitmusik 和Patorale 两个品种经辐射育种方法获得[11],但鲜见有关铁线莲辐射育种具体操作方法的研究报道。 本研究以威灵仙种子为试验材料,比较不同剂量137Cs-γ 射线辐射对威灵仙种子萌发、幼苗茎、叶表型变异和生理指标的影响,旨在筛选适宜的辐射剂量,为促进我国观赏铁线莲品种自主培育及种质资源开发提供参考。
1 材料与方法
1.1 试验材料
2016年12月从浙江农林大学铁线莲种质资源圃采集健康、成熟的威灵仙种子,去除尾部宿存花柱后用牛皮纸袋封装,4℃冰箱保存。 2017年1月在浙江省农业科学院作物和核技术利用研究所进行137Cs-γ 辐射处理,剂量率为1 Gy·min-1,分别设0(CK)、50、100、150、200 Gy 5 个辐射剂量水平,每个剂量辐射100 粒种子,重复3 次。 辐射后的种子在4℃冰箱放置1 周后,用200 mg·L-1赤霉素溶液浸泡24 h 后播种,以泥炭∶河沙(1 ∶1)作为栽培基质。 幼苗长出真叶后,移栽至泥炭土∶珍珠岩∶蛭石(3 ∶1 ∶1)的基质中,常规水肥管理。
1.2 试验方法
1.2.1 发芽势及发芽率统计 将两片子叶出土视为发芽标志,自第一颗种子发芽起,测定10 d 内萌发种子粒数和40 d 内萌发种子总数。 按照公式计算相关发芽指标:

1.2.2 半致死辐射剂量估算 参照周小梅等[12]的方法,用不同剂量辐射种子的相对发芽率拟合直线回归方程y=a + bx,计算半致死剂量(LD50)。
1.2.3 幼苗形态指标测定 观察成活植株形态变异情况,各处理随机选择30 株幼苗(150 Gy 和200 Gy 辐射剂量测定全部29 和14 株幼苗),在2018年5月10日、7月10 日和9月10 日分别测定不同剂量辐射处理幼苗分蘖数、株高、节数和节间长度,取平均值。2018年7月15-22 日,参照铁线莲属植物新品种DUS测试标准[10],对上述幼苗的叶形进行分类和统计,叶片位置以植株高度1/2 为界,分为“上部”和“下部”两部分。 按照以下公式计算各类叶形比例:

1.2.4 叶片相对叶绿素含量及叶绿素荧光参数测定 使用SPAD-520 叶绿素仪(日本柯尼卡美能达控股株式会社)和LI-6400 便携式光合测定仪(美国LICOR Biosciences 公司),各处理随机选择9 株幼苗,每株选3 片成熟、健康叶片,分别在2018年5月10 日、7月10 日和9月10 日上午8:30-10:30,测定叶片相对叶绿素含量和PSⅡ原初光能转化(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)、非光化学淬灭系数(non-photochemical quenching coefficient,NPQ)、光化学淬灭系数(photochemical quenching coefficient,qP)、电子传递速率(electronic transfer efficiency,ETR)等叶绿素荧光参数。
1.3 数据处理
采用SPSS 22.0 进行数据分析,采用Microsoft Office Excel 2007 制图。
2 结果与分析
2.1 137Cs-γ 射线辐射对威灵仙种子发芽的影响

图1 137Cs-γ 射线辐射对威灵仙种子萌发和成苗的影响Fig.1 Effects of 137Cs-γ radiation on the seed germination of Clematis chinensis
0 Gy 剂量处理组(CK)威灵仙种子播种后243 d开始萌发,50、100、150 和200 Gy 剂量处理组种子比CK 分别推迟5、10、10 和15 d 萌发。 由图1 可知,低剂量(50 Gy)辐射可提高威灵仙种子发芽势、发芽率和成苗率,较CK 分别增加3.47%,4.17%和6.37%;继续增加辐射剂量,上述各指标均显著下降。 将辐射剂量(y)与相应的种子相对发芽率(x)进行回归分析,得到回归方程y=-1.670x+193(R2=0.911),计算威灵仙种子辐射LD50为109.5 Gy。 就种子发芽而言,威灵仙种子辐射剂量以50 Gy 为宜。
2.2 137Cs-γ 射线辐射对威灵仙种子幼苗生长的影响
由表1 可知,137Cs-γ 射线辐射会造成幼苗分蘖数(图2)和节数减少,节间长度和株高(图3)减小。 CK幼苗分蘖数、株高、节数和节间长度变异系数均为最低,表明未经过辐射处理的威灵仙幼苗长势较整齐,且以上指标的最大值和最大均值都出现在CK 中,表明辐射对植株生长的影响主要表现为抑制作用。 辐射剂量对幼苗生长指标的变异效应影响表现为100 Gy >150 Gy>200 Gy>50 Gy>CK。
50 Gy 剂量处理组分蘖数变异植株比例在7月最高,达60%;100 Gy 剂量处理组分蘖数变异植株比例在7 和9月最高,均为80.00%;150 和200 Gy 剂量处理分蘖数变异植株比例在5-9月均达100%,表明较高的辐射剂量对幼苗分蘖能力的抑制效果显现更早,且抑制作用和持续性都较强。 随着辐射剂量的增加,植株矮化比例逐渐增加,200 Gy 剂量处理组植株矮化比例达到100%,表明高强度的辐射剂量对植株的矮化效应更明显。

图2 5月威灵仙幼苗分蘖情况Fig.2 Tiller status of Clematis chinensis seedlings in May

图3 5月威灵仙幼苗株高、节数、节间长度情况Fig.3 Seedling height,node number and internode length of Clematis chinensis seedlings in May

表1 137Cs-γ 射线辐射对威灵仙种子幼苗生长的影响Table 1 Effects of 137Cs-γ radiation on the growth of Clematis chinensis
2.3 137Cs-γ 射线辐射对威灵仙幼苗叶形变化的影响
依据铁线莲属植物新品种DUS 测试指南对叶片形态的分类方法,将CK 叶形分为6 种(图4、表2),全株均有分布。 其中卵形叶(1 号)占比最高,占CK 叶片总数的60.71%,卵状披针形叶(2 号)占比22.19%,具有以上2 种叶形的植株占比分别为93.33%和63.33%,卵形叶和卵状披针形叶为威灵仙基础叶形。按不同叶形的叶片数量由多到少排序为1 号>2 号>4号>5 号>6 号>3 号,可见威灵仙植株自然叶形丰富。
辐射处理后幼苗出现了7 种变异叶型(图5、表3)。其中倒卵状椭圆形叶(8 号)占比最高(14.03%),仅在50 Gy 剂量处理组出现。 圆形叶(13 号)占比最少(0.41%)。 按不同叶形的叶片数量由多到少排序为8号>9 号>11 号>7 号>10 号>12 号>13 号。 变异叶形中仅13 号叶边全缘,表明辐射对叶缘锯齿和叶裂的形成有促进作用。 7 号~13 号这7 种变异叶形主要出现在植株下部,其中7 号、11 号和12 号全部分布在植株下部,9 号在全株均有分布,其中有6.90%单独分布在植株下部,10 号变异叶有42.11%单独全部分布在下部,但8 号和13 号全部分布在植株上部。 表明多数叶形变异出现在幼苗生长的早期,随生长期推进,辐射积累效应有减弱的趋势。
50 Gy 剂量处理组出现的变异叶形种类最多,有5种,占50 Gy 总叶形种类的50%;150 Gy 剂量处理组出现的总叶形种类最多,有11 种,其中变异叶形种类占比45.45%;100 Gy 剂量处理组变异叶形占比为40%。上述结果表明,在50 ~150 Gy 剂量范围内,叶形较易受辐射影响发生变异。

图4 对照组(0 Gy)威灵仙叶形Fig.4 Leaf shape of Clematis chinensis without radiation dose treatment
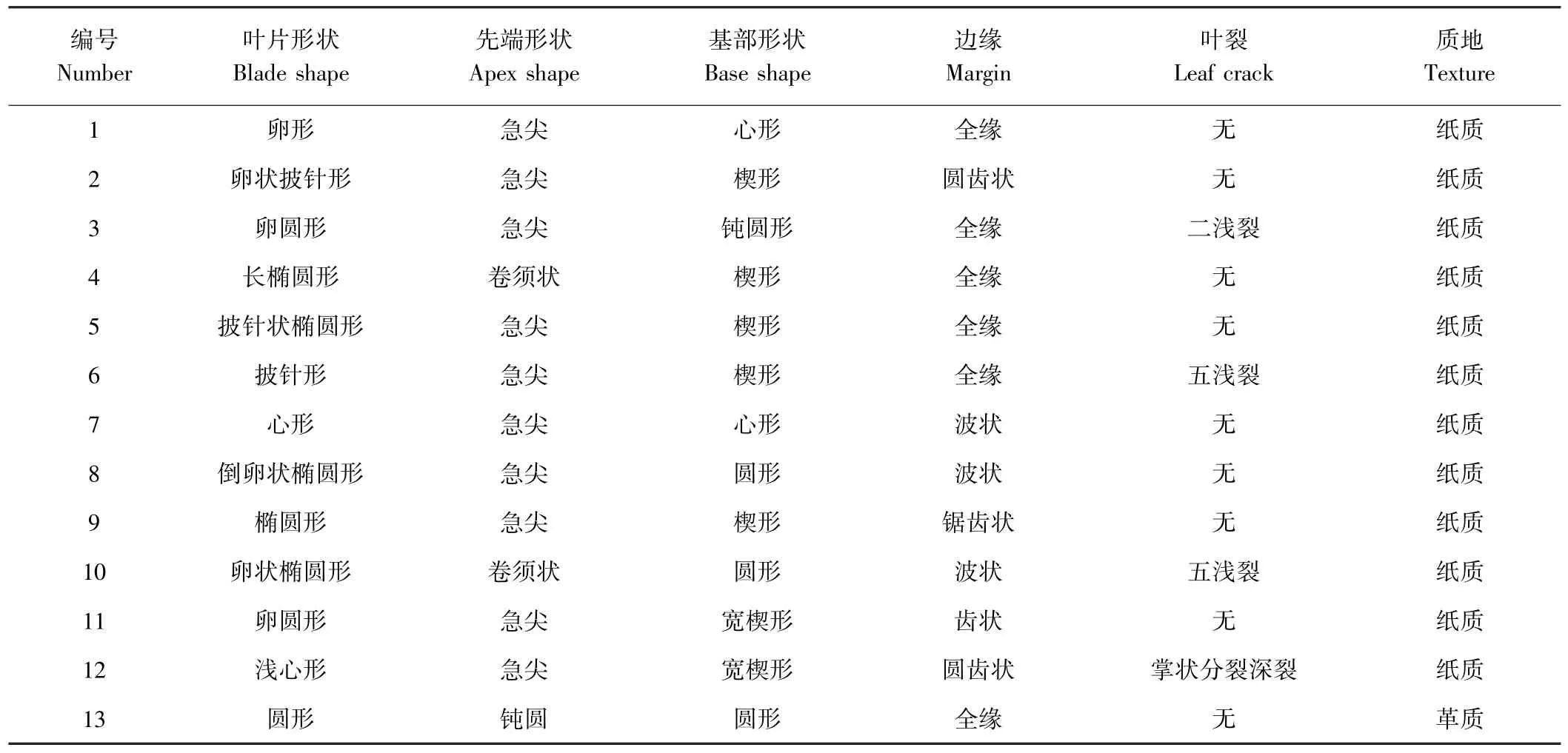
表2 威灵仙幼苗叶形Table 2 Leaf shape of Clematis chinensis seedling

表3 137Cs-γ 射线辐射对威灵仙幼苗叶形变化的影响Table 3 Effects of 137Cs-γ radiation on the leaf shape change of Clematis chinensis seedling
2.4 137Cs-γ 射线辐射对威灵仙幼苗相对叶绿素含量的影响
相对叶绿素含量是定量描述叶片绿色度的指标[13],其值高,植物能捕获更多光能进行光合作用[14]。 由图6 可知,50 ~200 Gy 剂量范围内,随着辐射剂量的增加,威灵仙幼苗叶片的相对叶绿素含量呈下降趋势,辐射剂量高于50 Gy 时,叶绿素合成受到抑制。 此外,叶片的相对叶绿素含量随生育期推进整体呈增加趋势,但其增幅随辐射剂量增加而减小,50、100、150 和200 Gy 剂量处理组叶片相对叶绿素含量在9月分别较5月增加了2.41%、2.41%、1.12%和0。上述结果表明,随着植株生长,137Cs-γ 射线辐射对叶绿素合成的抑制效应呈消减趋势。

图5 辐射(50、100、150 Gy)处理后威灵仙变异叶形Fig.5 New leaf shape of Clematis chinensis after 50、100、150 Gy radiation dose treatment

图6 137Cs-γ 射线辐射对威灵仙种子幼苗相对叶绿素含量的影响Fig.6 Effect of 137Cs-γ radiation on the relative chlorophyll content of Clematis chinensis
2.5 137Cs-γ 射线辐射对威灵仙幼苗叶绿素荧光的影响
由表4 可知,随着生长期的推移,各辐射处理的威灵仙幼苗Fv/Fm、Fv/Fo和ETR 均呈上升趋势,表明植株光合能力在持续恢复和提升。 5、7 和9月的数据均值显示,50 Gy 剂量辐射幼苗的Fv/Fm和ETR 较CK分别增加了0.99%和3.46%,辐射剂量继续增加(100~200 Gy),两指标总体呈下降趋势,说明低辐射剂量(50 Gy)对威灵仙PSⅡ利用光合的能力具有促进作用,而更高剂量(100~200 Gy)辐射下威灵仙的光合作用受到抑制。 NPQ 随着辐射剂量的增加总体呈下降趋势,相邻处理中50 ~100 Gy 剂量辐射的降幅最大,达到14.11%;qP 在50 ~200 Gy 剂量范围内随辐射剂量增加而增大,200 Gy 剂量辐射较CK 增加8.68%,表明在辐射导致的光抑制下,威灵仙会通过热耗散的形式进行自我保护。 综合各项指标认为,50 Gy 剂量辐射可以促进威灵仙利用光能,更高剂量辐射会使威灵仙光合作用受到抑制。
3 讨论
LD50是指植物受到辐射后导致半数植物死亡的辐射剂量,辐射临界剂量(LD40)是指受辐射植物存活率为对照组40%的辐射剂量[15],是确定该植物(器官)适宜辐射剂量的重要参考标准。 目前LD50没有统一的测定方法,通常采用种子发芽率、幼苗成活率、营养和生殖生长的抑制程度等指标来确定[16]。 适宜辐射剂量的选取也无统一标准,主要以植株成活率和有益变异性状出现的概率进行综合考量。 如熊云海等[17]将相对出苗率测算得到的LD50作为桂花辐射育种的适宜剂量;李奎等[18]用γ 射线照射黄牡丹种子后,发现经40~60 Gy 处理的苗高是对照组的50%,因此将40~60 Gy 作为适宜剂量范围;蔡春菊等[19]测定得到毛竹的LD40为175 Gy,毛竹对照组生长量70%的辐射剂量在100 Gy 左右,于是取100 ~175 Gy 为最适剂量范围。 根据前人研究结果,本研究依据LD50兼顾变异比例和生长情况,来确定威灵仙的适宜剂量范围。本研究中,辐照剂量在50~100 Gy 之间时,茎、叶变异比例高,且生长和光合系统运作状况良好,此外,根据相对发芽率测定的威灵仙LD50为109.5 Gy,因此将威灵仙的适宜剂量范围选为50~109.5 Gy。

表4 137Cs-γ 射线辐射对威灵仙幼苗叶绿素荧光参数的影响Table 4 Effects of 137Cs-γ radiation on chlorophyⅡfluorescence parameters of Clematis chinensis
辐射导致植物叶片形态变异较丰富,主要为单/复叶类型改变、叶缘改变和叶色改变。 如波士顿蕨在高剂量60Co-γ 射线辐射下叶片从一回羽状复叶突变为二回羽状复叶[20];板栗接穗经10 ~40 Gy 剂量辐射后叶片面积增大,且叶片出现卷曲[21];海州常山经15 ~45 Gy 剂量辐射后叶面出现缺绿和叶斑[22];铁线莲属植物叶片形态多样,且一个种类同时具有多种叶形的情况十分普遍[23]。 本研究中137Cs-γ 射线辐射诱导共得到7 种变异叶形,主要表现在叶缘、叶基部、叶先端和质地,说明辐射对威灵仙叶形变异效应显著,对新品种选育具有积极作用。 但辐射对威灵仙花型、花色等观赏性状的影响还有待进一步深入探究。
叶绿素是捕获光能的物质基础,其含量在一定程度上可反映植物同化物质的能力[24]。60Co-γ 射线辐射使越南油茶璠龙无性系叶绿素含量降低[25],廖安红等[26]和桂仁意等[27]也证明60Co-γ 和137Cs-γ 射线辐射会破坏叶片光合色素含量,刺梨和毛竹幼苗的叶绿素含量均随着辐射剂量增加而下降。 本研究中,与对照相比,威灵仙经50 Gy 剂量辐射后其相对叶绿素含量并未出现显著降低,表明低剂量辐射对叶绿素合成影响不显著,而高于100 Gy 剂量的辐射会增加对叶绿素合成的抑制作用,这与白刺花幼苗受辐照后类似[28]。 叶绿素荧光与光合作用中各个反应过程紧密相关,逆境对光合作用各过程的影响都可通过植物体内叶绿素荧光诱导动力学变化反映出来[29],同时叶绿素荧光参数更能反映植物“内在性”的特点[30]。 多花野牡丹Fv/Fm和Fv/Fo在0 ~55 Gy 剂量范围内均随辐照剂量的增加呈现先上升后下降的趋势[31],表明PSⅡ光能转化率先上升后下降,下降的原因可能是辐射导致PSⅡ反应中心运转情况不良,这与本试验中威灵仙受辐照后表现相同。 但何岭等[22]对海州常山进行60Co-γ 辐射,发现Fv/Fm和Fv/Fo均随辐射剂量的增加而降低,这可能是由于研究的材料有差别,或者试验的辐射剂量不同。 qP 可以反映植物光合活性,电子传递活性越大,qP 越大。 辐射处理增加了葡萄叶片的qP[32],本研究也得到相似结果,威灵仙受辐射后qP较对照组也有所增加。 结合各叶绿素荧光参数的表现,低剂量辐射可能会刺激植物中光合酶类的活性,使各指标表现提升,但过高的辐射剂量会破坏植物光合反应中心,导致植物生长受阻。
4 结论
本研究利用137Cs-γ 射线辐射威灵仙种子,发现50~200 Gy 剂量范围内,低剂量(50 Gy)辐射会促进威灵仙种子发芽,幼苗成活和生长;辐射剂量高于50 Gy 时威灵仙种子受损,成活率下降,幼苗光合能力减弱,生长受到抑制。 50~109.5 Gy 剂量范围内,威灵仙幼苗茎、叶变异比例较高,矮化现象明显,植株光合生理状态与长势较好,可作为威灵仙辐射育种适宜辐射剂量。
猜你喜欢
中国民族民间医药(2022年1期)2022-02-19 02:57:20
保健与生活(2022年4期)2022-02-13 10:43:05
中学生数理化·七年级数学人教版(2020年12期)2021-01-18 06:57:46
中学生数理化·七年级数学人教版(2019年12期)2019-05-21 02:53:50
中成药(2017年3期)2017-05-17 06:09:16
首都食品与医药(2014年3期)2014-10-19 03:04:42
语文教学与研究(读写天地)(2014年2期)2014-05-26 12:25:03
安徽医药(2014年4期)2014-03-20 13:12:36
语文教学与研究(2014年6期)2014-03-12 06:06:11
湖南农业科学(2014年10期)2014-02-27 14:29:40
