
补光时间及光质对温室甜椒矿质元素吸收与分配的影响
2020-07-03 08:00:32段青青张自坤常培培张禄祺王静静贺洪军
核农学报 2020年8期
段青青 张自坤 常培培 张禄祺 王静静 贺洪军
(德州市农业科学研究院,山东 德州 253015)
光是植物进行光合作用的主要能量来源,是诱发植物生长和结构分化的环境信号之一[1]。 光通过光强、光质和光周期调节植物细胞、组织、器官的形态建成、生长和分化,进而影响植物的生长发育。 光合有效辐射中红光(600~700 nm)和蓝光(400~460 nm)两个波段是能够被叶片吸收进行光合作用的主要光谱,且叶绿素吸收红光的比例大于蓝光[2],因此在进行人工补光时,通常红光的占比大于蓝光[3]。 与高压钠灯、荧光灯等人工光源相比,发光二极管(light emitting diode,LED)具有体积小、寿命长、能效高、光谱可调、环境友好等特点,已被广泛应用于园艺生产。 研究表明,LED 补光除了能够促进园艺作物的生长、增强植株光合作用、提高产量、改善产品品质[4-8],还能影响植物体内营养元素的积累及分配,进而调控植物的生理代谢及生长发育[9]。 甜椒(Capsium annuumL.)是我国设施栽培的主要蔬菜种类之一,冬春季节设施内的低温弱光环境往往造成甜椒生长发育受阻,采用LED 补光能够促进甜椒植株的生长,提高甜椒产量并改善甜椒的果实品质[10-11]。 矿质元素吸收积累量的增加有助于促进植物的生长,进而提高产品品质[12-13]。 但关于LED 补光对甜椒植株营养元素积累及分配的研究较少[14-15]。 高比例红光能增加果实的产量,而少量蓝光能够提高果实的商品性[16],甜椒结果期人工补光,以红光为主添加少量的蓝光可以同时提高果实的产量和品质[17-18]。 本试验选择以红光为主的不同红蓝比例的光质进行补光,以日光温室甜椒奥黛丽为试材,通过分析补光光质及补光时间对甜椒营养元素积累与分配的影响,探究补光调控的营养吸收机制,旨在为温室甜椒栽培的设施光调控提供一定的理论参考。
1 材料与方法
1.1 试验材料
试验于2017年11月至2018年1月在德州市农业科学研究院平原科技创新园15 号日光温室进行。供试甜椒品种为奥黛丽,购于山东安信种苗股份有限公司,采用穴盘基质育苗,育苗基质为草炭∶珍珠岩∶蛭石(体积比7 ∶3 ∶1),于2017年8月24 日(苗龄45 d)定植于日光温室内。
试验光源为定制LED 植物补光灯,购于惠州可道科技股份有限公司,灯具为长方形,尺寸长×宽×高为380 mm×280 mm×180 mm,单灯功率100 W,每盏灯由高亮度的红色(R,630 nm)和蓝色(B,460 nm)灯珠组成。
1.2 试验设计
试验采用二因素随机区组设计,因素A 为补光光质,设红光和蓝光组合R ∶B=2 ∶1(2R1B)、R ∶B=4 ∶1(4R1B)和R ∶B=8 ∶1(8R1B)3 个光质处理;因素B 为补光时间,设补光2 h(18:00-20:00)、补光4 h(18:00-22:00)和补光8 h(18:00-02:00)3 个补光时间;以不补光处理为对照(CK),共计10 个处理。 每个处理设3 次重复,每个重复为一个小区,小区面积为26 m2(南北6.5 m×东西4 m),每小区栽种90 株,小区随机排列。 补光灯悬吊于温室内,每小区4 支灯,在小区内安装间距为南北2 m,东西1.3 m,不同小区间用遮光布进行隔离。
甜椒在红蓝光下的光补偿点在45 μmol·m-2·s-1左右[19],且暗期较低光照强度补光即可对植株的生长发育产生影响[20-22],因此本试验补光光强设为55 μmol·m-2·s-1,通过调节光源与植物顶端的垂直距离,使各处理到达幼苗冠层的光量子通量密度相等。 光强及光谱测定采用PLA-20 植物光照分析仪(杭州远方光电信息股份有限公司),各处理光质光谱分布见图1,补光时间由定时器控制。 根据植株生长高度适时调整光源位置。
甜椒苗于2017年8月24 日定植于温室,前期光照强,未进行补光,随着温室内光照减弱,补光自11月1 日开始,此时植株处于初果期,株高50±2 cm,茎粗10±1 mm,叶片(叶长大于2 cm)数55±3 片,补光至2018年1月30 日结束,补光周期90 d,之后进行相关指标的测定,均设3 次重复。
1.3 测定项目与方法
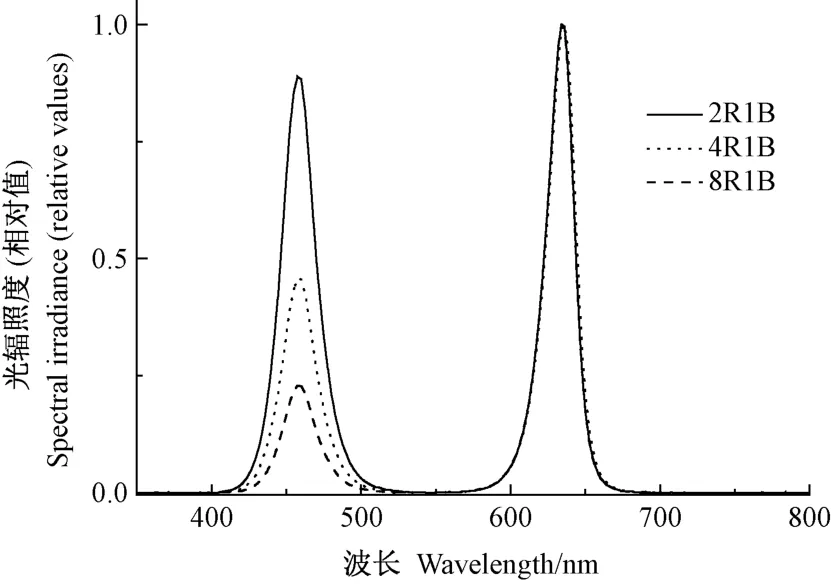
图1 不同光质的光谱分布图Fig.1 Spectral distribution graph of different light qualities
每处理选取5 株甜椒植株,分离地上部(茎、叶、果)和地下部(根),烘干后测定各部分干重,称重后的样品粉碎,过80 目筛备用。 采用凯氏定氮法测全氮(nitrogen,N) 含量,采用钼锑抗比色法测全磷(phosphorus,P) 含 量,采用火焰光度法测全钾(potassium,K)含量,采用原子吸收光谱法测定钙(calcium,Ca)、镁(magnesium,Mg)、铜(copper,Cu)、锌(zinc,Zn)含量[23]。 根据各器官的干物质量及各养分含量计算单株N、P、K、Ca、Mg、Cu、Zn 积累量。
1.4 数据分析
采用Execl 2010 软件进行数据处理,采用IBM SPSSStatistics 21.0(SPSS Inc,美国)统计软件进行方差分析,其中,双因素方差分析通过主体间效应检验对两因素间的交互作用进行验证,差异显著性分析采用Duncan 法进行检验(P<0.05),采用Origin8.1 软件作图。
2 结果与分析
2.1 补光时间及光质对甜椒干物质积累量的影响
补光时间及光质对甜椒的全株干物质积累量存在显著影响,除了光质2R1B 补光2 h 的干物质积累量与CK 无显著差异外,其余处理均显著高于CK(图2),即补光显著促进了甜椒干物质的积累。 同一补光时间下,干物质积累量随着红光比例的增大而升高;同一光质下,除2R1B 补光2 h 和4 h 的干物质积累量存在显著差异外,不同补光时间的干物质积累量均无显著差异。 双因素方差分析结果显示,补光时间对甜椒干物质积累量的影响不显著(F=1.674,P>0.05),而光质对甜椒干物质积累量影响显著(F=99.456,P<0.05),且光质与补光时间对甜椒干物质积累量存在显著的交互作用(F=11.041,P<0.05)。
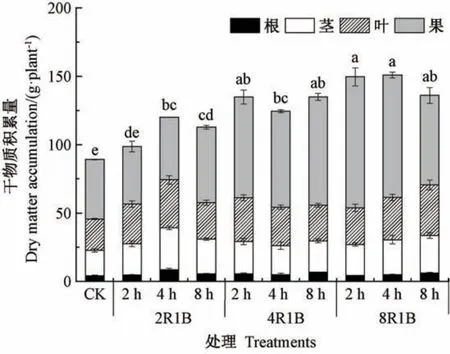
图2 补光时间及光质对甜椒干物质积累量的影响Fig.2 Effects of supplemental light and durations of illumination on dry matter accumulationof single sweet pepper plant
2.2 补光时间及光质对甜椒N 吸收积累与分配的影响
元素吸收积累量是元素含量与干物质积累量的乘积,因此干物质积累量和元素含量共同影响元素积累量的变化。 由表1 可知,甜椒N 积累总量与干物质积累量的变化趋势大致相似,即补光显著增加了甜椒N的总积累量。 不同的是,光质2R1B 补光2 h 的N 积累总量显著高于CK,而二者的干物质积累量无显著差异,主要是因为光质2R1B 补光2 h 的N 含量高于CK。 2R1B、4R1B、8R1B 3 种光质的N 积累总量分别为3.02、3.24、3.03 g·plant-1,相互之间差异不显著;但补光时间显著影响N 的积累,补光8、4 h 的N 积累总量分别较补光2 h 高18.02%和9.89%,且光质与补光时间对甜椒N 积累总量存在显著的交互作用(F=8.385,P<0.05)。
光质对N 在甜椒根、茎、叶、果实中的积累量无显著影响,补光时间显著影响N 在甜椒根、茎、果实中的积累,且光质与补光时间对N 在甜椒根、茎、叶、果实中的积累存在显著的交互作用。 补光2 h,N 在茎中的积累量高于补光4、8 h;而N 在果实中的积累量低于补光4、8 h。 从分配比例看,N 在甜椒植株各器官的分配比例从高到低依次为果实>叶>茎>根,可能是因为补光时期正是甜椒的结果期,此时N 开始向生殖器官中转移[24]。 补光2 h 有利于N 向甜椒根、茎、叶中分配,补光4、8 h 有利于N 向甜椒果实中分配。
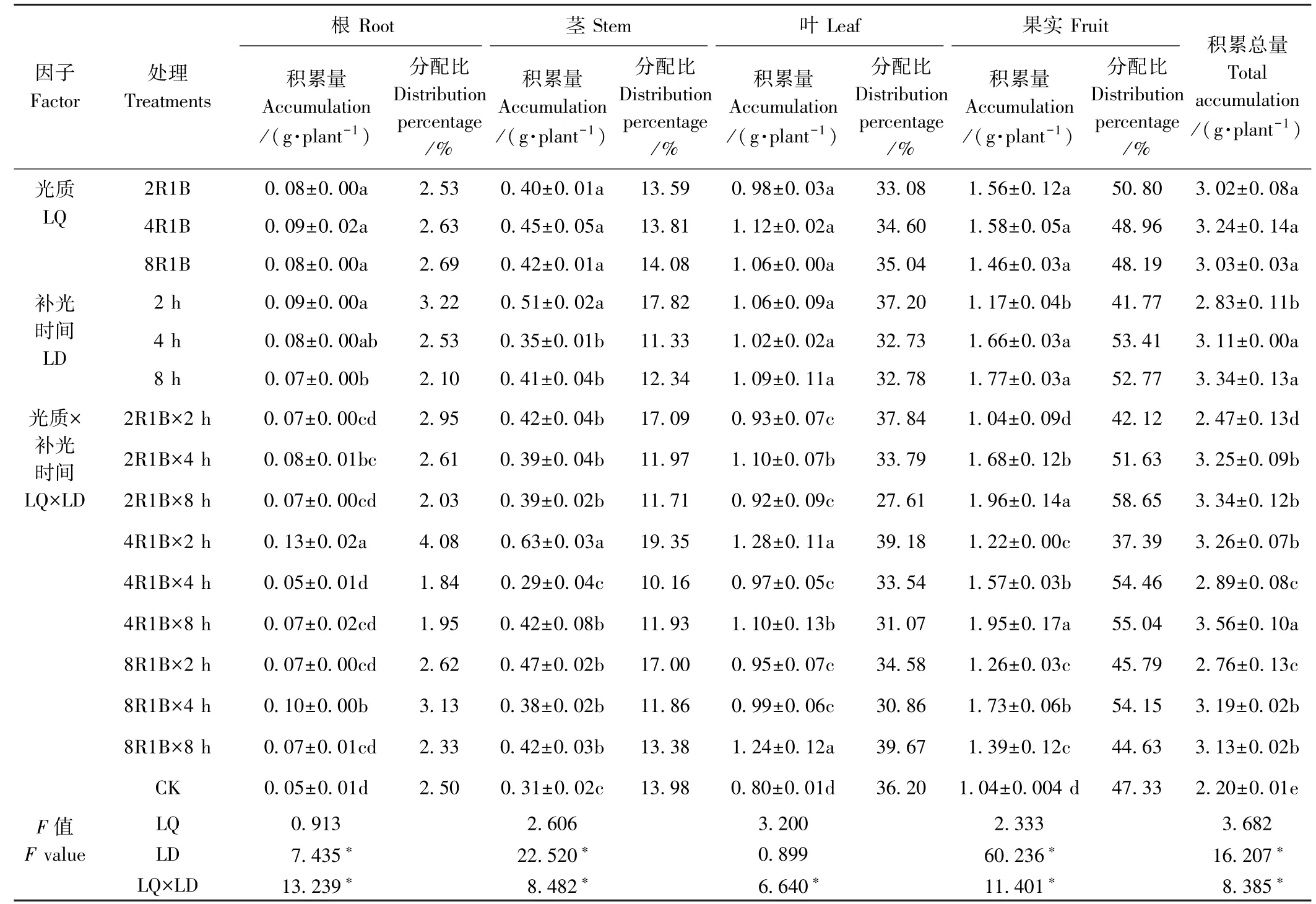
表1 补光时间及光质对甜椒N 积累与分配的影响Table 1 Effects of supplemental light and durations of illumination on N accumulation and distribution of sweet pepper
2.3 补光时间及光质对甜椒P 积累与分配的影响
光质和补光时间对P 在甜椒各器官积累量影响的双因素方差分析如表2 所示。 P 积累总量的变化趋势与干物质积累量的变化趋势相似,即除了光质2R1B补光2 h 的P 积累总量与CK 无显著差异外,其余补光处理的P 积累总量均显著高于CK。 2R1B、4R1B、8R1B 3 种光质P 的积累总量分别为0.36、0.37、0.35 g·plant-1,光质对甜椒P 积累总量影响不显著;但补光时间对P 积累总量影响显著,补光8 h 的P 积累总量为0.42 g·plant-1,分别较补光2、4 h 高40.00%和16.67%,且光质与补光时间对P 积累总量的影响存在显著的交互作用(F=7.536,P<0.05)。
光质及补光时间对P 在甜椒根和茎中的积累无显著影响,但光质对P 在叶中的积累影响显著,光质4R1B 叶中P 的积累量最高,显著高于光质2R1B;光质对P 在甜椒果实中的积累影响不显著,但补光时间显著影响了P 在果实中的积累,且补光8 h 果实的P积累量最高,显著高于补光4、2 h。 光质与补光时间对P 在甜椒叶和果实中的积累存在显著的交互作用。 P在甜椒各器官中的分配比例从高到低依次为果实>叶>茎>根,补光2 h 较CK 增加了P 在根、茎、叶中的分配比,降低了P 在果实中的分配比;补光4、8 h P 在果实中的分配比明显高于CK。
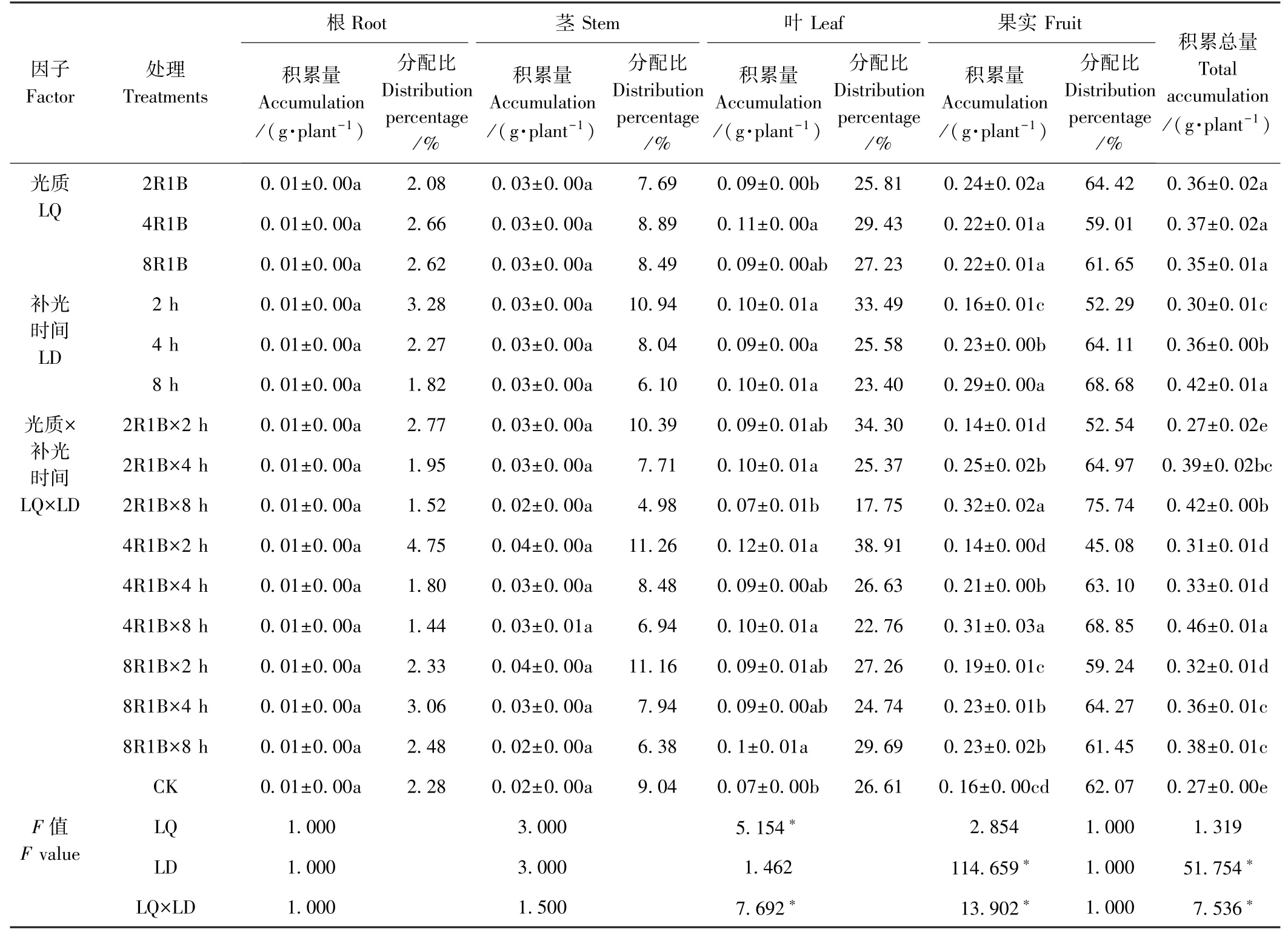
表2 补光时间及光质对甜椒P 积累与分配的影响Table 2 Effects of supplemental light and durations of illumination on P accumulation and distribution of sweet pepper
2.4 补光时间及光质对甜椒K 积累与分配的影响
由表3 可知,K 积累总量的变化趋势与干物质积累量的变化趋势大体一致,即光质2R1B 补光2 h 的K积累总量与CK 无显著差异,其余处理的K 积累总量均显著高于CK;不同的是光质4R1B 补光2 h 的干物质积累总量显著高于CK,但K 积累总量与CK 无显著差异,这主要因为光质4R1B 补光2 h 果实、茎、叶中的K 含量低于CK。 2R1B、4R1B、8R1B 3 种光质K 的积累总量分别为4.22、4.30、4.19 g·plant-1,双因素方差分析结果显示,光质对甜椒K 积累总量的影响不显著;但补光时间显著影响甜椒的K 积累总量,随着补光时间的延长,K 积累总量逐渐升高,补光8 h 的K 积累总量显著高于补光2、4 h,分别高34.73% 和11.09%。 光质与补光时间对甜椒K 积累总量的交互作用不显著(F=2.768,P>0.05)。
光质对K 在甜椒根、叶、果实中的积累量无显著影响,但对茎中K 的积累量影响显著,光质8R1B 茎中K 的积累量显著高于光质2R1B 和4R1B;补光时间对茎和果实中K 积累量的影响显著,补光8 h 茎和果实中K 的积累量显著高于补光2、4 h。 光质和补光时间对甜椒根和果实中的K 积累量具有显著的交互作用。另外,K 在茎中的积累量明显高于N 和P,这可能与K参与植物茎秆维管束发育有关[25]。 K 在甜椒各器官中的分配比例从高到低整体为果实>叶>茎>根,补光2 h 的K 在甜椒根、茎、叶中的分配比例高于CK,但不利于K 向甜椒果实中的分配;补光4 h 和8 h 的K 在甜椒果实中的分配比例高于CK。

表3 补光时间及光质对甜椒K 积累与分配的影响Table 3 Effects of supplemental light and durations of illumination on K accumulation and distribution of sweet pepper
2.5 补光时间及光质对甜椒Ca 积累与分配的影响
由表4 可知,Ca 积累总量的变化趋势与干物质积累趋势大致相似,但也有所不同。 光质2R1B 补光2 h Ca 积累总量与CK 无显著差异,其余处理均高于CK,不同的是光质4R1B 补光4 h 的干物质积累量显著高于CK,而其Ca 积累总量与CK 无显著差异,主要是该处理果实和根中的Ca 含量低于CK,尤其是果实中的Ca 含量显著低于CK 引起的。 双因素方差分析结果显示,光质和补光时间显著影响Ca 在甜椒中的积累总量,且二者对Ca 积累总量有显著的交互作用(F=16.286,P<0.05)。 光质8R1B 的Ca 积累总量显著高于光质2R1B 和4R1B,分别高11.72%和18.25%。 补光8、4 h 的Ca 积累总量分别为4.89 和4.74 g·plant-1,二者无显著差异,但均显著高于补光2 h,分别高32.52%和28.46%。
Ca 作为一种信号调节物质,能够保持植物体内的生理平衡[26]。 光质对Ca 在甜椒根、茎、叶、果实中的积累量具有显著影响,光质2R1B 果实中Ca 的积累量最高,光质4R1B 叶中Ca 的积累量最高,光质8R1B 根和茎中的Ca 积累量最高。 补光时间也显著影响Ca在甜椒茎、叶、果实中的积累,补光8 h 茎和果实中Ca的积累量最高,显著高于CK,补光2、4 h 叶中Ca 的积累量较高,显著高于CK。 但是补光4、8 h 果实中Ca的积累量无显著差异,且光质和补光时间对Ca 在甜椒根、茎、叶、果中的积累有显著的交互作用。 与N、P、K 在植株中的分配不同,Ca 主要分布在甜椒的茎和叶中,补光2 h 不利于Ca 向果实中分配,其分配比明显低于CK,而补光4、8 h 的Ca 在果实中的分配比明显高于CK。
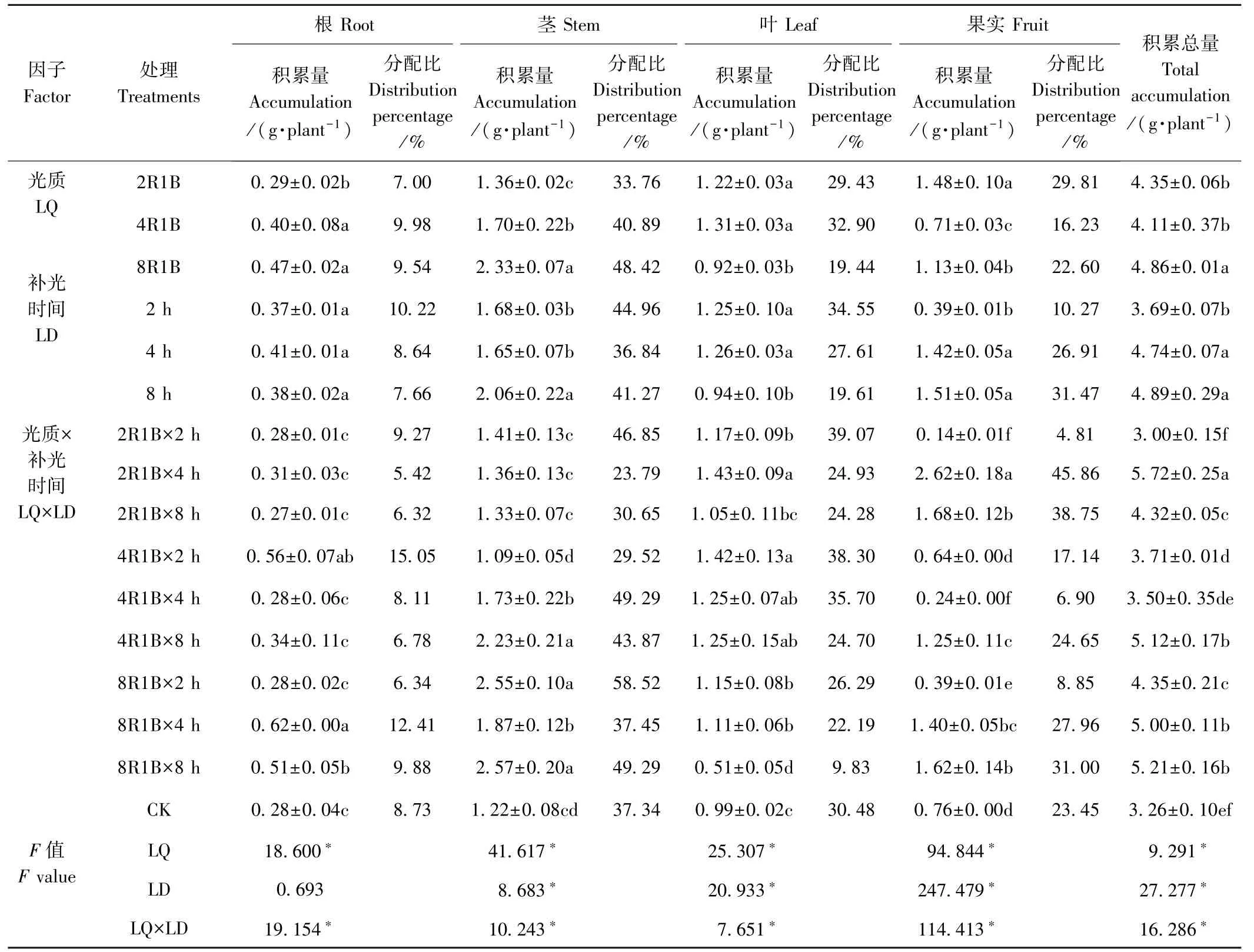
表4 补光时间及光质对甜椒Ca 积累与分配的影响Table 4 Effects of supplemental light and durations of illumination on Ca accumulation and distribution of sweet pepper
2.6 补光时间及光质对甜椒Mg 积累与分配的影响
由表5 可知,Mg 积累总量的变化趋势与干物质积累量的变化趋势大致相同,即补光能显著增加甜椒中Mg 的积累量,不同的是,光质2R1B 补光2 h 的干物质积累量与CK 无显著差异,但Mg 的积累总量显著高于CK,这是由Mg 在根、茎、叶中的含量高于CK 引起的。双因素方差分析结果显示,光质对Mg 的积累总量无显著影响,2R1B、4R1B、8R1B 3 种光质的Mg 积累总量分别为0.51、0.52、0.52 g·plant-1,相互之间差异不显著;但补光时间显著影响Mg 在甜椒中的积累,补光8 h Mg 的积累总量显著高于补光2、4 h,分别高28.89%和11.54%。 光质和补光时间对Mg 积累总量存在显著的互作效应(F=5.081,P<0.05)。
光质对Mg 在甜椒根、叶中的积累量无显著影响,但显著影响了Mg 在甜椒茎和果实中的积累,光质2R1B 果实中Mg 的积累量最高,而光质4R1B 和8R1B茎中Mg 的积累量显著高于2R1B。 补光时间显著影响Mg 在叶和果实中的积累,补光8 h 叶和果实中Mg的积累量最高,显著高于补光2 h,但补光8、4 h 叶和果实中Mg 的积累量无显著差异。 光质和补光时间对Mg 在甜椒根、叶、果实中的积累存在显著的互作效应。Mg 在甜椒植株中的分配以叶中最高,其次是果实、茎和根,这可能与Mg 是叶绿素的主要组成部分有关[26]。 补光2 h 的Mg 在果实中的分配比明显低于CK;补光4、8 h 的Mg 在果实中的分配比高于CK。
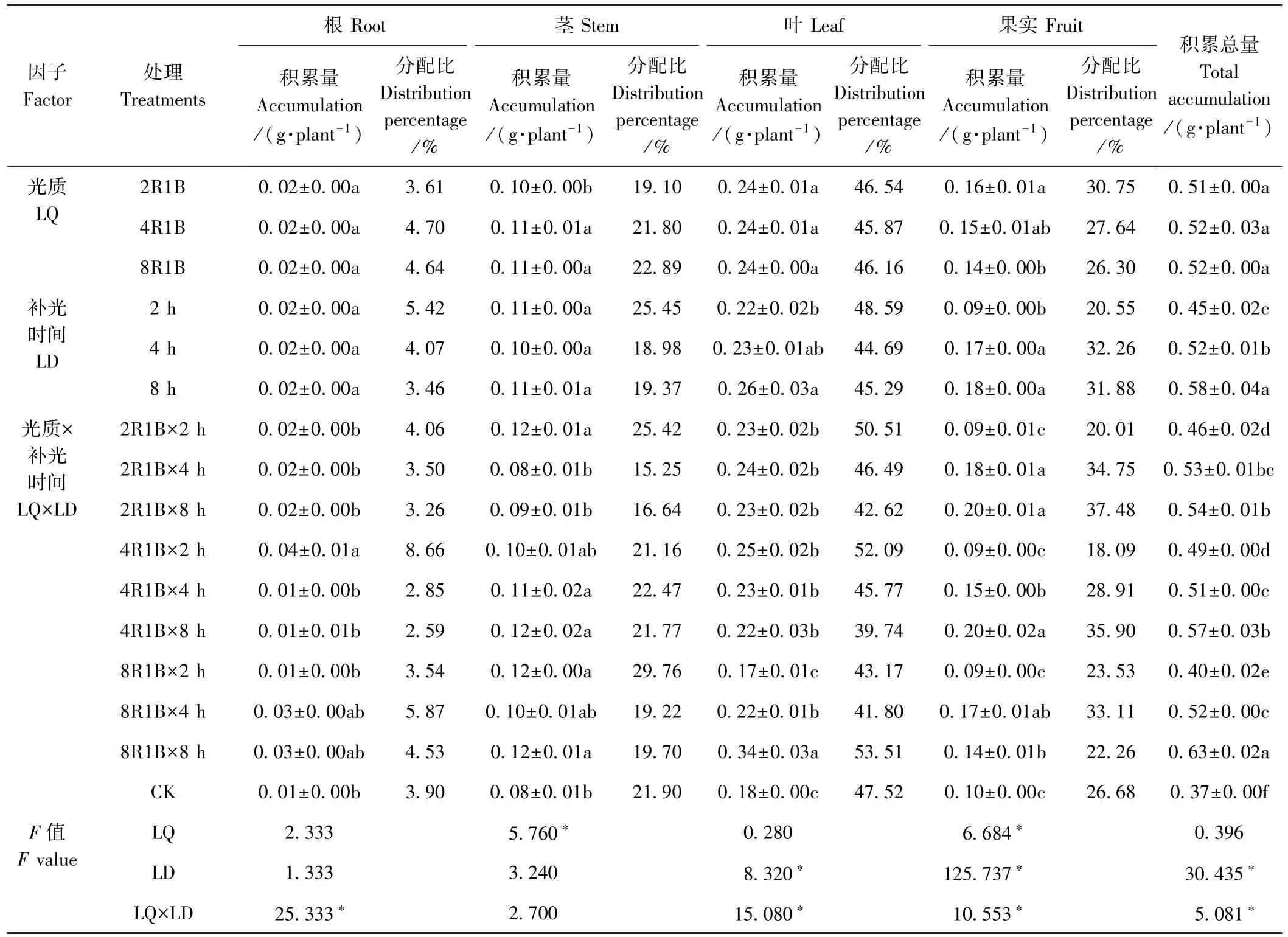
表5 补光时间及光质对甜椒Mg 积累与分配的影响Table 5 Effects of supplemental light and durations of illumination on Mg accumulation and distribution of sweet pepper
2.7 补光时间及光质对甜椒Mn 积累与分配的影响
由表6 可知,Mn 积累总量的变化趋势与干物质积累量的变化趋势相似,即补光能显著促进Mn 在甜椒植株中的积累,不同的是光质2R1B 补光2 h 的干物质积累总量与CK 无显著差异,但Mn 积累量显著高于CK,这是由该处理Mn 在叶中的含量高于CK 引起的。2R1B、4R1B、8R1B 3 种光质的Mn 积累总量分别为3.32、3.55、3.49 mg·plant-1,双因素方差分析结果显示,光质对Mn 的积累总量无显著影响;但补光时间显著影响Mn 的积累总量,补光2、4、8 h 的Mn 积累总量分别为3.19、3.55、3.61 mg·plant-1,补光4、8 h 的Mn积累总量无显著差异,但比补光2 h 分别高11.29%和13.17%。 光质与补光时间对Mn 在甜椒中的积累有显著的交互作用(F=21.881,P<0.05)。
光质对Mn 在甜椒根、茎、叶、果实中的积累量有显著影响,其中光质2R1B 果实中Mn 的积累量最高,光质4R1B 叶中Mn 的积累量最高,光质8R1B 茎和根中Mn 的积累量最高。 补光时间对根和叶中Mn 的积累量无显著影响,但显著影响茎和果实中Mn 的积累量,补光4 h 果实中Mn 的积累量显著高于补光2、8 h,而补光8 h 茎中Mn 的积累量显著高于补光2、4 h。 光质和补光时间对Mn 在甜椒根、茎、叶、果实中的积累有显著的交互作用。 Mn 在甜椒植株中的分配以叶中最多,其次是果实、茎和根,这可能与Mn 参与叶绿体的形态建成及促进光合电子传递的功能有关[27]。 补光时间影响了Mn 在甜椒各器官分配,与CK 相比,补光不利于Mn 在茎中的分配;补光2 h 提高了Mn 在叶中的分配比;补光4 h 的Mn 在果实中分配比明显高于CK。

表6 补光时间及光质对甜椒Mn 积累与分配的影响Table 6 Effects of supplemental light and durations of illumination on Mn accumulation and distribution of sweet pepper
2.8 补光时间及光质对甜椒Zn 积累与分配的影响
由表7 可知,Zn 积累总量的变化趋势与干物质积累量的变化趋势不同,但光质2R1B 补光2、8 h、光质8R1B 补光2 h 的Zn 积累量均显著低于CK,这是由于这些处理茎、叶、果实中的Zn 含量低于CK,尤其是叶和果实中的Zn 含量明显低于CK 引起的。 双因素方差分析结果显示,光质显著影响Zn 的积累总量,2R1B、4R1B、8R1B 3 种光质的Zn 积累总量分别为1.65、1.96、2.00 mg·plant-1,后两者差异不显著,但分别比2R1B 高18.79%和21.21%。 补光时间也显著影响Zn 的积累,补光4 h 的Zn 积累总量最高,分别较补光8、2 h 显著高18.72%和45.10%。 光质与补光时间对Zn 在甜椒中的积累有显著的交互作用(F=25.492,P<0.05)。
光质对果实中Zn 的积累量无显著影响,但显著影响Zn 在甜椒根、茎、叶中的积累,其中光质8R1B 根和茎中Zn 的积累量最高,光质4R1B 叶中Zn 的积累量最高。 补光时间也显著影响Zn 在甜椒根、茎、叶、果实中的积累,其中补光8 h 根、茎、叶中Zn 的积累量最高,补光4 h 果实中Zn 的积累量最高。 光质和补光时间对甜椒根、茎、叶、果实中Zn 的积累有显著的交互作用。 补光时间对甜椒各器官中Zn 的分配影响较大,与CK 相比,补光2、8 h 提高了Zn 在甜椒根和茎中的分配比,降低了在果实中的分配比,而补光4 h 提高了Zn在果实中的分配比。
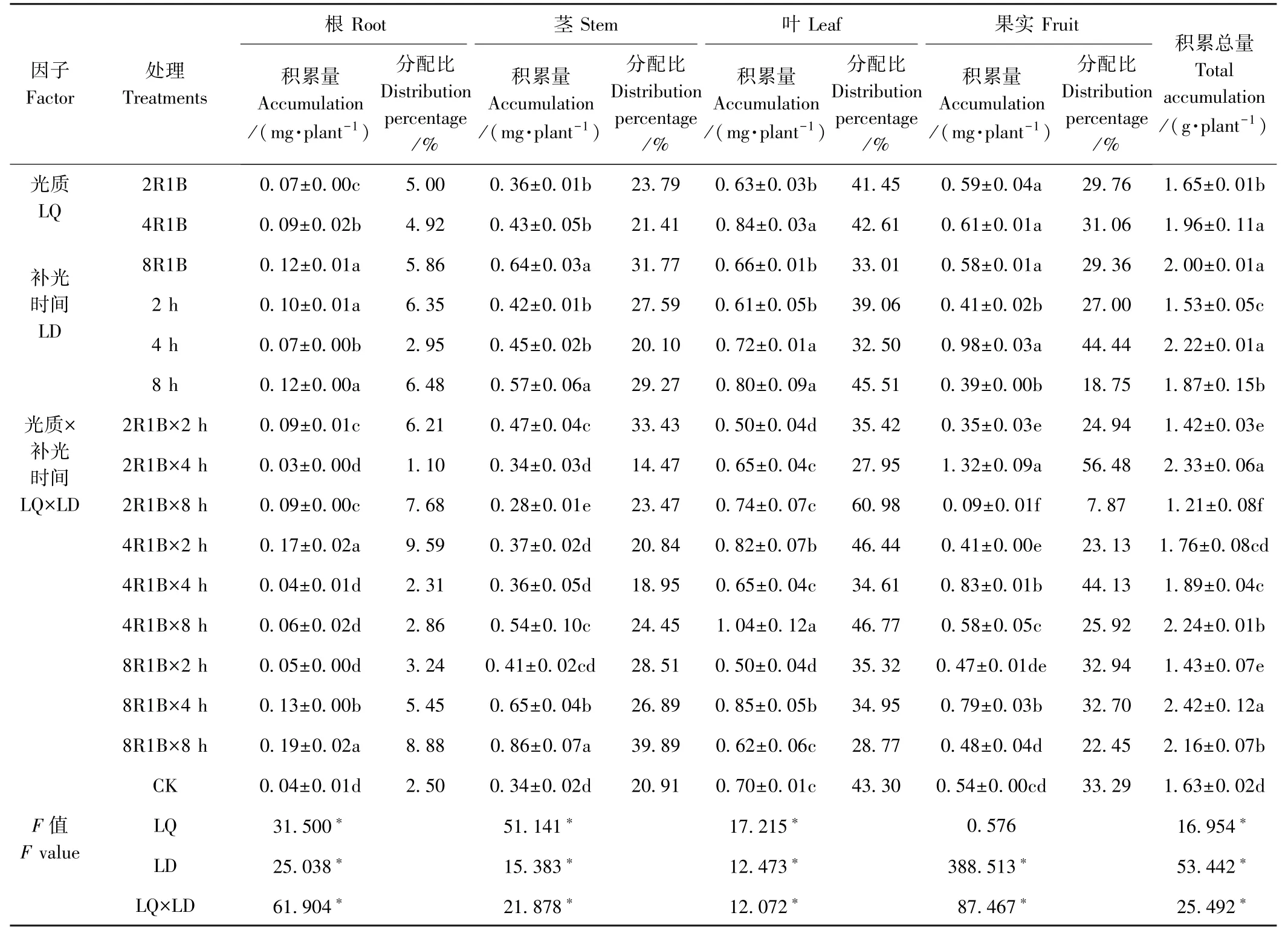
表7 补光时间及光质对甜椒Zn 积累与分配的影响Table 7 Effects of supplemental light and durations of illumination on Zn accumulation and distribution of sweet pepper
3 讨论
矿质元素在植物生长发育中起到非常重要的作用,可以直接或间接参与植物的生理生化及代谢过程[27]。 在甜椒整个生育期,结果期至成熟期的养分吸收量占整个生育期养分吸收总量的50%[28],甜椒结果期对矿质元素的需求量从高到低依次为K>N>Ca>Mg>P[29]。 前人研究表明,随着生育期的推进,甜椒的干物质积累量不断增加,对矿质元素的吸收量也逐渐增加,因此元素的积累量不断升高,但不同元素种类的积累量变化趋势不一致[30-31]。 本试验结果显示,甜椒的干物质积累与元素积累量的变化趋势大致相似。 与CK 相比,LED 补光能在显著增加甜椒干物质积累的同时,显著增加N、P、K、Ca、Mg、Mn 在甜椒全株的积累,尤其是补光8 h 的N、P、K、Ca、Mg、Mn 积累总量分别较CK 提高51.82%、55.56%、47.55%、50.00%、56.76%、36.04%,这与前人的研究结果相符[32-33]。 可见,通过LED 补光能够提高甜椒对矿质元素的吸收积累能力,可以考虑将其作为调控甜椒养分吸收的一种农艺措施。
红蓝组合光质对营养元素积累的影响因植物种类、光质配比、补光时间等因素存在明显差异[34-35]。前人研究表明,不同红蓝比例对生菜和甜罗勒中N 的积累量无显著影响[36-37],而红蓝比80 ∶20 提高水培生菜中Ca、Zn 含量的效果优于其他光质配比[38]。 本试验中,2R1B、4R1B、8R1B 3 种光质对N、P、K、Mg 和Mn的积累总量无显著影响,但光质8R1B 的Ca 和Zn 积累总量高于2R1B、4R1B,尤其显著高于2R1B,这与前人研究结果相似[38]。 此外,适宜的补光时间能够增加矿质元素的积累,但对不同元素种类的影响也不尽一致。 有研究表明,补光12 h 能显著增加黄瓜幼苗中的N、Ca、Mg、Zn、Cu、Fe 的积累量[39],而补光4 h 显著提高了生菜中K 和Ca 的吸收量[40]。 本试验中,补光时间显著影响了N、P、K、Ca、Mg、Mn、Zn 的积累量,补光4 h 的Zn 积累总量最高;补光8 h 的P、K、Mg 积累总量最高;补光4、8 h 的N、Ca、Mn 积累量无显著差异,但均显著高于补光2 h。 这与前人研究不完全一致,可能是植物种类、补光时间、光照强度等不同造成的,还需要进一步研究。 另外,光质和补光时间对甜椒元素积累量的影响存在显著的交互作用。 本试验结果显示,2R1B 补光4 h 的Ca 和Zn 积累总量最高,4R1B 补光8 h 的N、P、K 积累量最高,8R1B 补光8 h 的Mg 和Mn 积累总量最高。 元素含量的变化与植株的生理功能密切相关[41-43],本试验中LED 补光引起的元素积累量的变化,对甜椒的光合特性、色素含量、渗透调节能力及抗病性等生理功能的影响还需要进一步研究。
光质及补光时间不仅影响植株矿质元素的总积累量,对矿质元素在植株各器官中的积累分配也存在显著影响[38,44]。 本试验中,光质和补光时间对元素在各器官中的积累分配存在显著的交互作用。 其中,2R1B补光4、8 h 的元素在果实中积累分配较高,4R1B 补光2 h 的元素在根、叶中积累分配较高,而8R1B 补光8 h的元素在茎中分配较高。 可见,适当增加红光比例有利于元素向茎、叶等营养器官中分配转移,而适当增加蓝光比例利于元素向果实中分配转移,这与前人的研究结果一致[12,35]。 此外,前人研究表明,延长补光时间能够增加矿质元素的积累,尤其是地上部矿质元素积累量[39]。 本试验中,与CK 相比,补光2 h 降低了N、P、K、Ca、Mg、Mn 在果实中的分配比;补光4 h 增加了N、P、K、Ca、Mg、Mn 在果实中的分配比,且补光8、4 h 果实中的元素积累量均显著高于补光2 h,这与前人研究结论是一致的[39]。 由此可见,补光2 h 不利于矿质元素向果实中的分配转移,而补光4、8 h 能够促进矿质元素向果实中的分配转移,而矿质元素向果实中分配转移率的提高,有利于形成高产[45]。
4 结论
光质与补光时间能显著影响矿质元素在甜椒中的积累与分配,且二者存在交互作用。 LED 补光能显著促进N、P、K、Ca、Mg 和Mn 在甜椒全株的积累。2R1B、4R1B、8R1B 3 种光质对甜椒N、P、K、Mg、Mn 积累总量无显著影响。 补光时间显著影响矿质元素的积累分配,补光2 h 降低了N、P、Mg、Mn 在果实中的分配比,补光4 h 提高了N、P、K、Mg、Mn 在果实中的积累总量和分配比。 综上,光质对甜椒矿质元素积累量的影响较小,LED 补光4 h 在调控甜椒营养积累与分配上表现最优。 本试验结果为温室栽培甜椒补光调控的营养吸收机制提供了理论参考。 但本试验仅研究了结果期补光对元素积累与分配的影响,不同生育期补光试验还有待进一步开展研究。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:08:50
四川林业科技(2022年1期)2022-03-22 07:48:32
上海农业学报(2022年1期)2022-03-08 00:09:12
电子制作(2019年19期)2019-11-23 08:41:52
今日农业(2019年16期)2019-01-03 11:39:20
幸福(2018年15期)2018-06-02 07:55:55
幸福·健康版(2018年5期)2018-05-09 07:06:06
中国交通信息化(2017年8期)2017-06-06 07:16:44
浙江农业学报(2017年1期)2017-05-17 06:13:36
饮食与健康·下旬刊(2016年5期)2016-11-28 10:55:11
