
神经营养因子在运动调控帕金森病理机制中的作用研究进展
2020-07-01 03:07:42贾芳芳张宪亮
体育科学 2020年3期
余 锋,贾芳芳,张宪亮
(1.淮阴师范学院 体育学院,江苏 淮安 223300;2.苏州大学 体育学院,江苏 苏州 215021;3.山东大学 体育学院,山东 济南 250061)
帕金森病(Parkinson’s disease,PD)是一种与衰老相关、以神经细胞分子生物学功能紊乱为特征、具有独特病因学基础的神经退行性疾病。PD的病理基础是中脑黑质致密部(Substantia nigra pars compacta,SNpc)基底神经节多巴胺(dopamine,DA)能神经元的丢失、神经递质DA减少而导致的渐进性神经系统功能障碍(Beal,2010)。DA能神经元的缺失导致DA的产生与消耗失衡,故PD患者表现出静息性肌肉震颤、运动性肌力缺失、运动迟缓、姿势僵硬不稳、步态异常等临床病理学特征(Lenka et al.,2015)。目前,PD的病因尚不明确,关于PD的病理机制存在几种学说,如神经炎症致病学说、氧化应激致病学说、线粒体功能障碍致病学说等(Ryan et al.,2015)。
研究发现,运动在降低神经系统疾病,如阿尔茨海默症(Alzheimer’s disease,AD)(Corsi et al.,2016)和 PD(Xu et al.,2010)等的患病风险中发挥重要作用,可延缓这些神经退行性疾病的恶化(Speelman et al.,2011),促进PD患者肢体运动能力的恢复(Takano et al.,2014),延缓PD大鼠纹状体DA丢失(刘晓莉等,2018),抑制纹状体神经元的过度兴奋,改善PD大鼠运动能力(刘晓莉等,2014),但其中的生物学机制尚不明确。大量动物实验和临床检验发现,神经营养因子(neurotrophic factors,NFs)的缺乏是动物或人患PD的重要原因,因为NFs的表达增加可促进PD的运动能力增强和病症缓解。随着运动疗法在PD干预中的效果日益凸显,运动干预与PD获益的潜在机制引起学界的关注。运动干预上调NFs的表达进而改善PD病症,可能是其中的机制之一(Hsueh et al.,2018;Speck et al.,2018;Tuon et al.,2014)。NFs可能在运动干预DA能神经元存活的过程中发挥关键作用,但运动如何通过调节NFs介导PD病理?其内在机制是什么?本文对近年来运动介导NFs调控PD的动物实验和人体试验研究进行分析,探讨运动介导NFs改善PD的可能调节机制,以期明晰上述疑问。
1 NFs的神经保护功能
NFs是一类在中枢和外周均发挥生物学功能的蛋白分子。在细胞核内,NFs可诱导基因编码,进而促进神经元的存活和增殖(Ibanez,2007)。脑内的NFs种类繁多,包括脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)、神经营养因子-3/4/5(neurotrophin 3/4/5,NT-3/4/5)、神经生长因子(nerve growth factor,NGF)、胶质细胞源性神经营养因子(glial cell-derived neurotrophic factor,GDNF)、星形胶质细胞源性神经营养因子(mesencephalic astrocyte derived neurotrophic Factor,MANF)、胰岛素样生长因子-1(insulin-like growth factor 1,IGF-1)和血管内皮生长因子(vascular endothelial growth factor,VEGF)等各NFs家族成员。NFs在神经元的生长、发育及凋亡等过程中发挥重要作用,可促进神经元的增殖、分化、成熟、突触发生等生理属性(Kirik et al.,2004;Sullivan et al.,2011)。NFs促进神经元存活的功能,被认为是治疗PD等神经退行性疾病的有效途径(Kirik et al.,2004)。有研究认为(Kirik et al.,2004;Vijender et al.,2015),NFs为大分子蛋白,不易跨越血脑屏障(brain-blood barrier,BBB)由血液进入脑脊液,临床治疗中人工合成的外源性NFs难以发挥其神经保护作用。但也有研究发现(Pan et al.,1998;Wu et al.,2014),NFs可以通过BBB介导血与脑中营养成分的动态平衡。
PD病理可能与多种NFs的缺失有关,但目前运动科学研究中涉及的NFs种类并不多,其中,BDNF、GDNF、IGF-1和VEGF,尤其BDNF是运动干预PD动物实验和人体试验研究的焦点。研究发现,合理的运动干预可促进BDNF(Hsueh et al.,2018;Tuon et al.,2014;Wu et al.,2011)、GDNF(Lau et al.,2011;Speck et al.,2018)、VEGF(Al-Jarrah et al.,2010)和 IGF-1(Maass et al.,2016)等的表达,改善神经系统的营养供应,促进神经元的存活、再生与分化,缓解神经退行性变化等,增强神经保护作用,改善PD病理,降低PD患病风险。
1.1 脑源性神经营养因子(BDNF)
BDNF是NFs家族的重要成员,参与神经元的生存、分化、生长、损伤后修复,以及新突触发生、神经突起的改建、突触传递等结构和功能的可塑性(Rothman et al.,2013)。BDNF广泛分布于中枢神经系统,尤其在海马区大量表达(Mutlu et al.,2016),亦表达于黑质纹状体中,参与调节DA能神经元的存活和功能维持(Lau et al.,2011)。研究发现,在众多NFs中,BDNF与DA能神经元的存活和分化关系最为紧密(Lau et al.,2011;Tajiri et al.,2010;Tuon et al.,2012;Wu et al.,2011)。体外培养的啮齿类动物神经细胞实验(Rak et al.,2014;Santos et al.,2016)证实,BDNF表达增加可促进黑质纹状体的神经元存活。它对酪氨酸激酶受体具有较强的亲和力,可作为第一信号分子结合到细胞表面,参与调节cAMP反应元件结合蛋白(cAMP-response element binding protein,CREB)的合成。CREB是一种细胞转录因子,可结合到环腺苷酸反应元件(CRE)上特定的DNA序列,参与相关基因转录的调节。BDNF可通过N-甲基-D-天冬氨酸受体(N-methyl-D-aspartic acid receptor,NMDAR)调节钙调蛋白激酶(calcium/calmodulin protein kinase II,CAMKII),激活丝裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)级联反应,进而诱导突触可塑性相关基因表达(Platenik et al.,2000)。此外,BDNF还可通过其受体TrkB 激活 MAPK,通过促进突触素 I(Synapsin I,Syn I)(Hong et al.,2015)和CREB(Mutlu,2016)的磷酸化调节突触可塑性。
BDNF在运动介导的神经保护机制中发挥重要作用。它可介导运动改善神经可塑性、脑损伤后的神经元恢复及学习记忆等重要脑功能(Sajadi et al.,2017)。运动可有效促进脊髓损伤大鼠脊髓内BDNF的表达,以及其后肢运动功能的恢复(贺晓玉 等,2014)。Lan等(2014)研究发现,跑台运动可促进脑梗死大鼠皮质BDNF和5-羟色胺(5-hydroxytryptamine,5-HT)及其受体亚型1A(5-HT1A)的表达增加。有氧游泳运动可上调衰老大鼠海马BDNF的表达,改善其学习记忆能力(付燕等,2015)。人体试验(刘瑾彦等,2018)也发现,3个月的运动锻炼可促进60~69岁老年人外周血中BDNF水平的上调和认知功能的改善。体力活动量与机体安静状态下血液中的BDNF水平密切相关,经常参加体力活动的个体安静状态下血液中的BDNF水平低于体力活动少的个体(Nofuji et al.,2008)。这表明体力活动量较大的个体,其安静状态下外周血中的BDNF较低可能与其穿过BBB入脑,发挥神经调节功能的利用率提高有关。综上研究表明,运动干预可通过增强脑和外周血中的BDNF表达,增强“血-脑”之间的BDNF传输,改善衰老或损伤的神经元功能,促进神经保护作用。
1.2 胶质细胞源性神经营养因子(GDNF)
GDNF来源于神经胶质细胞,参与了神经元的功能维持和神经损伤的修复(Enterria et al.,2016),可促进神经元的发育、存活、分化、凋亡和轴突发生,并能特异性保护DA能神经元,促进损伤后运动神经元的再生(Deng et al.,2013),对中枢和外周神经系统均具有保护作用。GDNF集中表达于儿茶酚胺类激素调控的脑区,如纹状体和丘脑(Pascual et al.,2008)。GDNF及其配体家族(GDNF family ligands,GFL)与DA能神经元的存活密切相关。GFL家族属于转化生长因子β(transforming growth factor,TGF-β)超级家族成员,在调控细胞迁移和分化过程中发挥作用(Hegarty et al.,2014)。GDNF具有神经保护和神经再生的双重神经营养功能。TGF-β作为GDNF信号的调制器,参与了细胞膜上GFL受体(GFRα)的迁移,并在细胞膜上与各自的配体相结合。GDNF配体和受体相关联,形成GFL-GFRα复合体,进而促进跨膜酪氨酸激酶(RET)二聚体的聚集。GDNF对GFRα-1有很强的亲和力,可形成GFL-GFRα-1复合体。聚集的RET也可反过来触发RET激酶分子的酪氨酸残基发生转磷酸作用,进而启动细胞内信号过程,引发相关细胞信号级联反应,如MAPK和磷酸肌醇3-激酶(phosphoinositide 3 kinase,PI3K)相关信号通路(Zhou et al.,2015),调控细胞增殖和转录过程,激活 CREB和蛋白激酶B(protein kinase B,Akt),在神经元的生长和存活中发挥重要作用。
GDNF也参与运动介导的神经保护,维持受损脊髓神经元的存活,促进轴突再生,抑制神经元的异常凋亡,促进神经损伤后的运动功能恢复。研究(蔡洪梅等,2016)发现,跑台运动可促进脊髓损伤大鼠脊神经中GDNF的分泌,减缓运动神经元的凋亡,增强轴突再生,改善脊髓损伤大鼠的运动功能。1个月的递增负荷运动干预可通过激活GDNF下游效应分子——细胞外信号调节激酶(extracellular regulated protein kinases,ERK),促进 GDNF的表达(Lau et al.,2011)。GDNF还可通过调节GFRα-1触发细胞信号级联,抑制小胶质细胞激活(Rocha et al.,2012)。4周的递增负荷游泳训练可促进小鼠海马GDNF和GFRα-1的表达(吴春燕 等,2011)。GDNF的表达增加,可能是机体在负荷训练增大时为避免神经元损伤的一种自我保护性反应。这表明GDNF和GFRα-1相关信号转导通路参与了随运动强度递增而产生的神经生物学调控过程,运动干预可通过促进NFs的表达来缓解神经损伤,促进细胞损伤的修复与再生。
1.3 胰岛素样生长因子(IGF-1)
IGF-1是一种受生长激素调节的多功能细胞增殖调控因子,具有多种生物学功能,包括维持机体能量代谢、调节血脂浓度和胰岛素活性(Street et al.,2013)等生命活动。除了参与调节躯体功能外,IGF-1在调节神经元的功能中也具有重要意义(Feng et al.,2015),可促进神经元的分化与存活。在成年人的神经系统中,IGF-1作为一种神经调质,参与突触可塑性、突触密度、神经传递和神经发生等过程的调节,尤其在 IGF-1受体(IGF-1 receptor,IGF-1R)集中表达的脑区发挥作用,可促进细胞外基质合成和神经元损伤的修复(Ayadi et al.,2016)。IGF-1R是一种酪氨酸激酶受体,主要表达于黑质DA能神经元和海马组织(Quesada et al.,2007),可通过触发细胞内相关信号转导途径介导IGF-1的多种生物学功能(Hernandez-Garzon et al.,2016)。IGF-1与脑高级功能的维持关系密切,参与学习记忆、缓解神经损伤和促进神经功能重塑等调控过程。在神经退行性病变过程中,IGF-1是重要的分子标志物,其水平的降低与衰老导致的脑功能下降以及神经退行性疾病(如PD和AD)相关。IGF-1可激活 PI3K/Akt-CREB等信号通路,调控促凋亡相关基因的转录(Liu et al.,2017)。脑内IGF-1受体被阻断可导致DA能神经元的死亡(Nikolić et al,2013)。
IGF-1也是运动介导神经保护功能的重要NFs。运动诱导外周血液中的IGF-1表达增多,是其发挥神经保护功能的重要方式之一。Cetinkaya等(2013)研究发现,规律的有氧运动可增加海马和血液中IGF-1的表达水平。运动还可增加脑对循环血液中的IGF-1摄取,预防脑的损伤,保护脑功能和改善认知(Maass et al.,2016)。IGF-1还可诱导神经元的增殖。Trejo等(2001)研究证明,运动可促进外周血中的IGF-1表达和脑对外周血中IGF-1的摄取,诱导成年大鼠海马新生神经元的增殖。利用IGF-1抑制剂阻断IGF-1的表达,发现运动对海马神经元的增殖促进作用被抑制。俞嘉玲等(2014)研究也证实,15天的短期自主运动可增强海马齿状回BDNF和IGF1的表达,促进成年小鼠海马的神经发生。这说明IGF-1在运动介导新神经元的发生和预防、缓解神经损伤、促进损伤神经组织修复的过程中发挥神经保护作用。
1.4 血管内皮生长因子(VEGF)
机体的内环境稳态依赖各级血管对氧和营养的转运,而大脑是对氧和营养物质需求极其旺盛的器官,氧或营养的缺乏可严重影响脑的高级功能发挥。促进脑的氧与营养供应可改善脑功能。运动可促进神经微血管的可塑性变化,其过程是通过促进脑内新生血管的形成和改善脑的血液循环来实现的,与运动对神经微血管发生相关因素的调节有关。VEGF是血管营养相关的生物因子,其功能是促进血管发生和受损血管的修复,在血流供应不足的区域恢复氧气供应(Yang et al.,2010;Zelzer et al.,2004)。VEGF家族成员由 VEGF-A、VEGF-B、VEGF-C、VEGF-D及胎盘生长因子(placental growth factor,PlGF)等构成(Ferrara et al.,1989)。其中,VEGF-A/B在对抗神经退行性疾病中发挥重要作用(Zechariah et al.,2013)。VEGF的异常表达及其功能障碍与炎症过程、PD和AD等神经退行性疾病有关(Yasuda et al.,2007)。VEGF参与了DA能神经元和神经胶质细胞的增殖与存活过程。研究发现,VEGF-A/B在PD大鼠中脑黑质DA能神经元中呈过表达状态(Falk et al.,2009)。VEGF家族成员可结合并激活3种不同类型的酪氨酸激酶受体,形成VEGF与酪氨酸激酶二聚体。细胞质中的酪氨酸激酶受体可进行自身磷酸化,触发MAPK、Akt和磷脂酶C(phospholipase C,PLC)等一系列的信号级联反应,促进血管新生和神经保护作用发挥(Huang et al.,2017)。
运动引起的机体氧耗增大,导致供氧不足及氧的利用能力下降,造成运动性缺氧,是诱导组织中VEGF表达增加的直接诱因(梁丽娟等,2005)。研究发现(Ding et al.,2006;Swain et al.,2003),运动可通过促进VEGF表达调控神经血管的生成和预防神经血管损伤,还可诱发脑部缺氧,促进VEGF的表达增加,继发性诱导衰老动物脑内新神经元的增殖,调控神经发生(李静等,2018)。Al-Jarrah等(2010)研究发现,有氧耐力运动可促进纹状体VEGF的表达,增加纹状体微血管密度,提高纹状体神经元的存活率。这表明,运动干预可通过诱发VEGF表达升高,刺激脑血管生成增加,改善脑组织氧和营养的供应环境,促进神经元的增殖或存活,介导神经保护,促进损伤神经组织的修复。4种NFs在运动介导神经保护中的可能机制如图1所示。
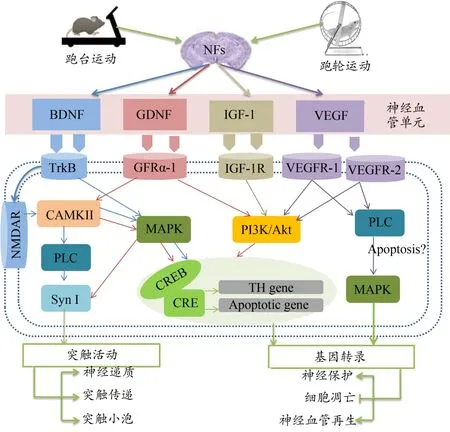
图1 NFs在运动介导神经保护中的可能机制Figure 1.Possible Mechanism of NFs in Exercise Mediating Neuroprotection
2 运动介导NFs干预PD的实验研究
随着生物医药技术的进步,PD的干预和治疗模式发生了重大变化,出现了诸如免疫疗法、基因疗法、干细胞疗法、脑深部电刺激和磁共振疗法以及运动疗法等新型干预模式。其中,运动疗法以费用经济、效应持久等优点,受到学界的广泛认可。在PD的运动干预中,NFs发挥了重要的神经保护作用。大量动物实验和人体试验均表明,运动干预可促进NFs的表达,维护DA能神经元的功能,改善PD。
2.1 运动介导NFs干预PD的动物实验
在PD的生物学机制研究中,动物实验发挥了极其重要的作用,选择合适的PD动物模型是研究得以深入的前提。常见的PD动物造模方式有两种:一是通过天然存在或人工合成的神经毒素造模;二是通过诱导PD相关基因的突变造模。前一种是近年来实验研究中较普遍采用的模式,其共同机制是通过神经毒素诱导DA能神经元的死亡(东惟玲 等,2014)。Zhou等(2017)研究发现,神经元内氨基酸受体的过度激活,会导致神经元的兴奋性毒性机制参与介导PD的神经元死亡,尤其是谷氨酸受体引起的神经毒性。常用的PD造模神经毒素,如1-甲基-4-苯基-1、2、3、6-四氢吡啶注射(1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine,MPTP)(Faherty et al.,2005;Gerecke et al.,2010)、6-羟多巴胺(6-hydroxydopamine,6-OHDA)(Hsueh et al.,2018;Speck et al.,2018)、脂多糖(lipopolysaccharide,LPS)(Kim et al.,2013;Wu et al.,2011)和鱼藤酮(rotenone)(Falk et al.,2009)等,均可导致啮齿类动物产生PD样病理特征,它们是当前PD动物实验研究中常用的模型诱导剂。
运动干预可缓解神经毒素诱导的PD模型鼠脑SNpc中DA缺失的症状。在PD大/小鼠的运动干预方案中,常采用自主跑轮运动和被动跑台运动两种模式(Ke et al.,2011),其中,跑台运动是采用较多的干预模式。研究发现,跑台运动和跑轮运动均可改善PD大/小鼠的病理症状,缓解DA能神经元的损伤,其机制与运动诱导的NFs表达增多有关(表1)。
综合表1中研究结果发现,短期(3~4周)、中期(6~8周)和长期(3 m~18周)的跑台运动、抗阻运动或自主运动等不同模式运动干预,均可促进PD模型鼠脑内的NFs表达,调控PD病理相关分子的水平,改善PD模型鼠的躯体运动能力。但不同模式运动对PD模型鼠的干预效应不存在明显差异,均可对PD病理发挥调控作用。
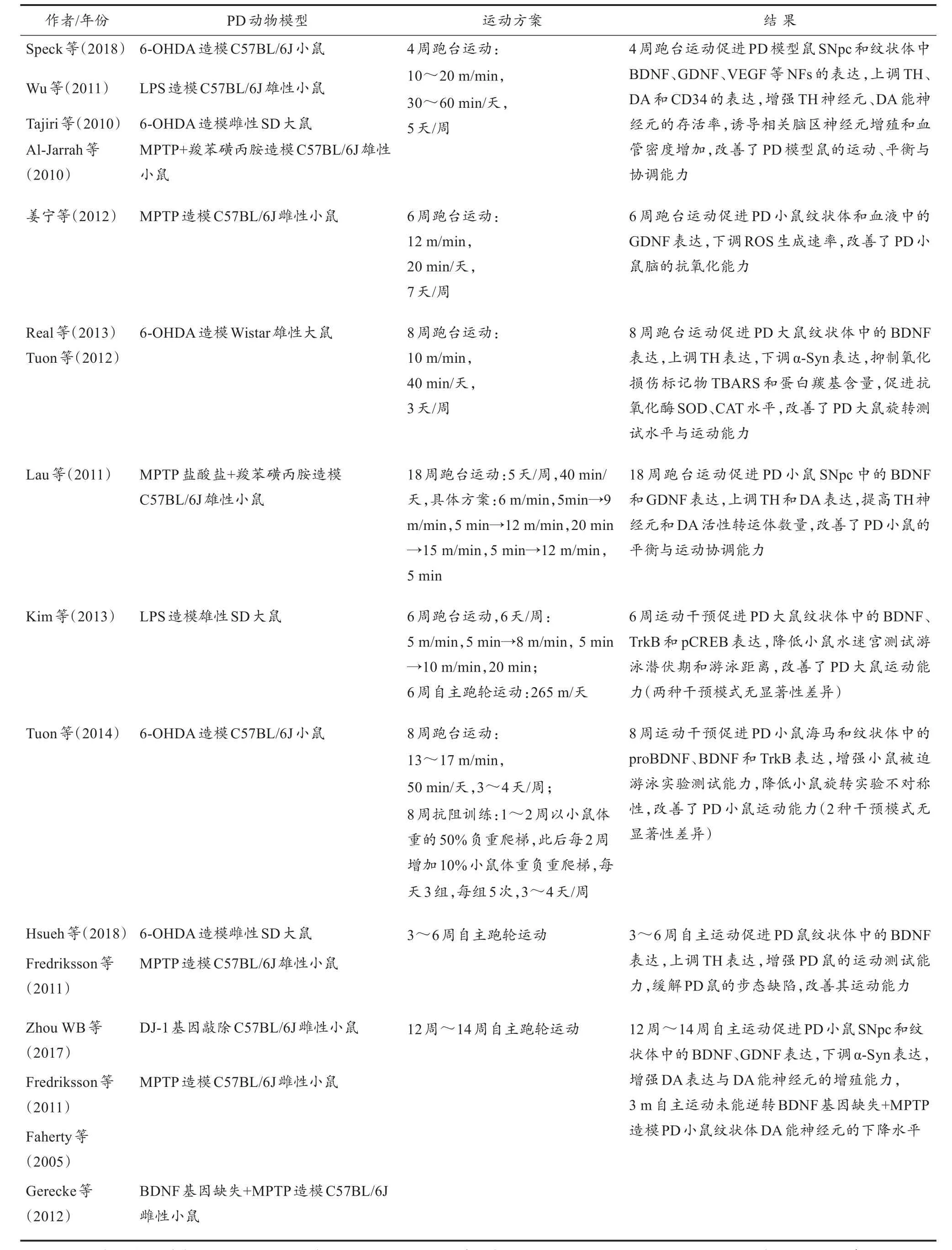
表1 运动干预PD动物模型的实验研究Table 1 Experimental Research of Exercise Intervention on PD Animal Models
2.2 运动介导NFs干预PD的人体试验
在运动干预PD的研究中,动物实验在探索疾病的生物学机制方面有其独特价值,是临床研究的重要依据。随着研究的深入,PD的人体试验逐渐开展起来。NFs在运动干预PD的人体研究中也发挥了重要作用。其中,BDNF是研究最多的NFs。研究发现,运动干预可通过促进PD患者血液中的BDNF表达,增强BDNF分子相关信号通路的靶向作用,改善PD患者的运动功能,促进PD病症的好转,为PD的早期预防提供重要参考。
在运动能力障碍PD的人体运动干预试验中,干预手段除经典的自行车运动(Marusiak et al.,2015;Zoladz et al.,2014)之外,新型干预模式如运动康复训练(motor rehabilitation training,MRT)(Angelucci et al.,2016)、强化运动康复治疗(intensive rehabilitation treatment,IRT)(Frazzitta et al.,2014)及其改进模式综合强化运动康复治疗(multidisciplinary intensive rehabilitation treatment,MIRT)(Fontanesi et al.,2016)是研究者常选且干预效果较好的模式,对改善PD患者的运动功能和缓解病症具有较好的借鉴价值。
测试量表是检测PD患者运动干预效果较直观的参考手段。常用的PD检测量表有PD统一评分量表(unified Parkinson’s disease rating scale,UPDRS)、贝格平衡量表(Berg balance scale,BBS)、PD残疾量表(Parkinson’s disease disability scale,PDDS)、蒙特利尔认知评估量表(Montreal cognitive assessment,MOCA)、简易智力状态检查量表(montgomery-asberg depression rating scale,MADRS)、PD患者睡眠状况评定量表(scale for outcomes in Parkinson’s disease-sleep,SCOPA-Sleep)、PD患者生活质量问卷(Parkinson’s disease questionnaire-39,PDQ-39)和步态调查问卷(freezing of gait questionnaire,FOGQ)等,以及PD相关运动能力测试,如6 min步行测试(6-minute walk test,6-MWT)和计时测试(time up and go test,TUG)等。研究发现,运动干预可通过促进BDNF的水平,改善PD患者相关量表和运动测试能力(表2)。
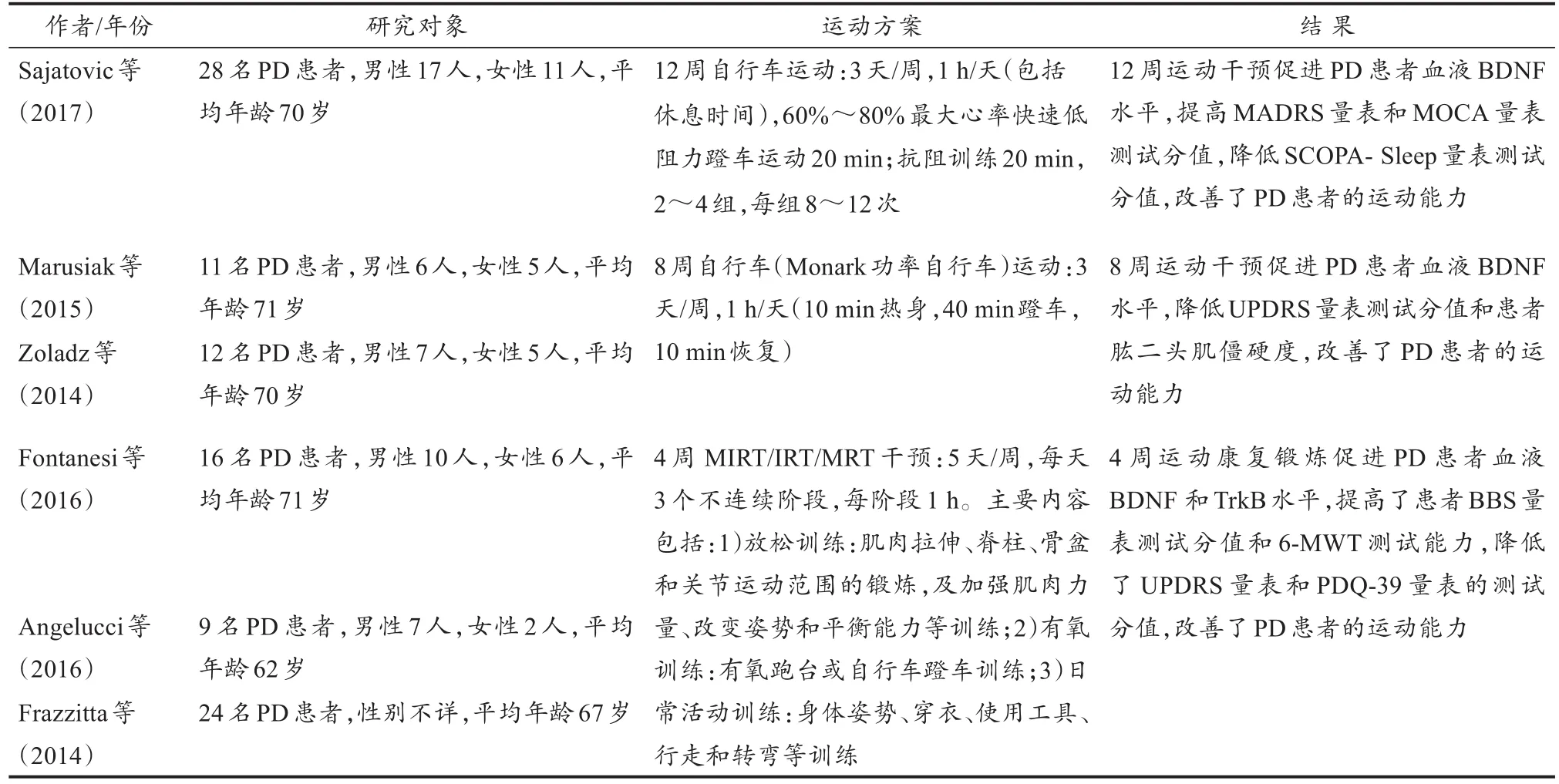
表2 运动干预PD的人体试验研究Table 2 Experimental Research of Exercise on Human PD
综合表2中的PD人体试验研究结果发现,8周和12周的自行车蹬车运动与4周的新型运动康复干预均可促进PD患者血液中的BDNF水平,改善其相关量表的测试分值,增强身体运动与生活自理能力。
3 NFs在运动介导PD病理中的可能机制
综合运动干预PD的动物实验和人体试验发现,运动干预可促进PD运动功能康复和病症改善,是预防和缓解PD的有效手段。NFs是参与中脑黑质—纹状体通路中DA能神经元存活和功能恢复的关键因子(Petzinger et al.,2007;Tajiri et al.,2010;Yoon et al.,2007),在运动干预诱导PD实验动物和PD患者运动行为能力改善过程中发挥了重要作用。运动干预可增加NFs的表达,促进DA能神经元的存活及相关信号分子的变化(Fredriksson et al.,2011;Gerecke et al.,2012;Griesbach et al.,2009;Wu et al.,2007,2011)。然而,运动干预介导NFs调节DA能神经元改善PD的机制尚不明晰。
3.1 运动可介导NFs调控TH干预PD
酪氨酸羟化酶(tyrosine hydroxylase,TH)集中表达于中脑、间脑、黑质及下丘脑等区域的儿茶酚胺能神经元和交感神经节及其支配的去甲肾上腺素能神经元,是介导与PD病理相关分子——DA合成过程中的重要限速酶。TH通过作用于酪氨酸,使其转化为L-二羟基苯丙氨酸,后者又通过L-氨基酸脱羧酶芳族酶转化为DA(Shulman et al.,2011)。TH活性的增加,可促进DA能神经元的存活(Qi et al.,2014)。在PD中,TH可诱导产生活性氧(reactive oxygen species,ROS),并易受ROS的攻击而失去活性。Sharma等(2016)通过大鼠黑质注射LPS诱导PD模型的研究发现,PD模型大鼠脑内总ROS量增加,进而导致TH的表达下降和DA的水平降低。因此,TH可能是调节神经功能正常的重要分子,其表达和功能的异常,是诱导神经退行性病变和导致PD的重要原因,也是PD干预与治疗的关键分子靶点之一。
运动可介导NFs调控TH的表达与活性,促进DA能神经元的发生和存活,调节神经可塑性,改善PD病理。研究发现(Hsueh et al.,2018;Lau et al.,2011;Real et al.,2013;Speck et al.,2018;Tajiri et al.,2010;Tuon et al.,2012;Wu et al.,2011),运动干预可通过促进 BDNF 和GDNF的表达,逆转6-OHDA或MPTP诱导的PD动物模型黑质纹状体中的TH失活,促进PD动物脑内的DA能神经元存活与功能恢复。这表明,运动可通过诱导PD脑内的NFs表达增加,促使TH活化,缓解DA能神经元的缺失或功能障碍。其机制可能与CREB有关。BDNF可结合并激活其受体酪氨酸激酶B(tyrosine kinase B,TrkB),进而触发BDNF下游的信号分子CREB。BDNF对TrkB受体的活化,是TH基因转录的调控因子之一(Fukuchi et al.,2010)。中枢TH的表达和活性受Ca2+、cAMP、糖皮质激素等多因子调节,其中,cAMP可通过CREB的磷酸化激活TH基因启动子来调节TH的表达(Nagatsu,2007)。在运动调节PD神经可塑性的机制中,CREB可直接参与调节TH的基因表达(Fukuchi et al.,2010)。因CREB是BDNF信号下游的重要分子,故其可介导BDNF与TH之间的信号传导。
3.2 运动可介导NFs调控NMDAR干预PD
DA能系统功能降低会进一步引起皮层-纹状体谷氨酸(glutamate,Glu)能通路的过度激活,Glu释放增加,诱发兴奋性神经毒性作用,进而导致DA/Glu失衡和基底神经节环路的功能紊乱(Schapira et al.,2014)。Glu是广泛分布于中枢神经系统的兴奋性神经递质之一,需要通过与其相应受体结合发挥功能。NMDAR是Glu的关键受体之一,是突触信息传递和突触可塑性变化的重要调节型受体(张凌韬 等,2018)。Davies等(2008)研究发现,PD发病时,神经元Glu释放增多,导致NMDAR的异常激活,促使神经元Ca2+内流增加,引起Ca2+超载,神经元损伤,即Glu的兴奋性毒性作用。Ferguson等(2014)实验证实,MPTP诱导的PD小鼠纹状体神经元外Glu浓度显著升高,NMDAR呈过度激活状态。
研究表明,BDNF可促进NMDAR的正常活化,缓解Glu通路的异常,降低Glu引起的兴奋性神经毒性。Black(1999)研究发现,BDNF可通过促进NMDAR的通道开放,进而增强突触后NMDA亚单位的磷酸化水平,激活NMDAR。孙彩霞等(2011)进一步证实,BDNF可通过增强突触后膜NMDAR的磷酸化,促进NMDAR的激活。运动可通过调节NFs的表达,促进NMDAR的正常生理性激活,抵抗Glu神经毒性,缓解Glu引起的PD病理。娄淑杰等(2007)实验发现,运动可增强实验大鼠脑内BDNF的表达,进而促进NMDAR的水平。运动还可通过NMDAR信号通路激活CREB,调节海马和纹状体的神经活动(Vaynman et al.,2003)。这说明,运动不仅能够直接调节NMDAR的水平,还可通过BDNF间接调控NMDAR的水平,以缓解Glu神经毒性,降低PD的风险。其原因可能与运动引起脑血流量增加,从而引起血—脑之间的BDNF和NMDAR穿梭效率提高有关(Vaynman et al.,2006)。
3.3 运动可介导NFs调控突触素干预PD
突触(synapse)是神经信息传导通路中神经元间信息传递的重要结构。突触传递效率随神经活动而变化的能力,即突触可塑性(synaptic plasticity),是神经系统结构和功能完整性的重要形式,参与脑认知功能的维持与发展。突触功能的异常和突触可塑性的损害,与PD和AD等神经退行性疾病密切相关(Kessels et al.,2009)。突触素(synapsin,Syn)是突触可塑性的重要分子标志,在神经信息处理中发挥重要作用。NFs特别是BDNF对Syn和突触可塑性的调节具有重要意义,可诱导轴突结构和功能变化(Cohen-Cory et al.,2010)。BDNF可以通过上调Syn的表达而改善突触可塑性(Gourley et al.,2013)。其中,SynI在NFs介导的突触可塑性中起关键作用。
运动可促进NFs增多,进而调节Syn的水平、介导突触结构和功能的可塑性,改善PD。运动干预可上调海马BDNF的表达,介导Syn增加,进而增强海马神经元的突触可塑性(Ferreira et al.,2011)。Vaynman等(2006)研究发现,在自主运动介导BDNF和SynI表达的实验中,通过TrkB-IgG模拟BDNF受体TrkB来选择性阻断BDNF的功能,则SynI的表达被抑制。这说明BDNF在运动调控SynI的表达中发挥关键作用。同时,IGF-1也可通过调节SynI的活性介导突触可塑性。Ploughman等(2005)研究发现,自主运动可促进脑缺血大鼠海马和皮层BDNF、IGF-1 与 Syn I的 表 达 。 IGF-1 可通过激活 CaMKII和MAPK信号通路,调节SynI的释放,改善突触功能(Ogundele et al.,2018)。由此可知,运动可通过促进脑内NFs介导Syn的水平,改进突触功能和突触可塑性。但目前缺少直接证据证明运动干预可通过调节NFs介导突触可塑性改善PD,未来的研究可将Syn作为运动干预NFs介导PD机制的分子靶点,以明晰Syn在运动干预PD过程中的确切机理。
3.4 运动可介导NFs调控α-突触核蛋白干预PD
α-突触核蛋白(α-synuclein,α-Syn)是一种与Syn结构不同、功能相反的PD致病性蛋白分子,集中表达于神经突触前,可调节神经元DA的释放。其所产生的神经毒性是导致DA能神经元死亡的诱因之一。α-Syn表达增多,可损伤培养中的原代神经元的轴突生长和分支(Koch et al.,2015),诱导人胚胎神经祖细胞的分化异常,导致神经元分化减少,神经胶质细胞分化增多(Zasso et al.,2018),影响正常神经功能。此外,α-Syn聚集可激活小胶质细胞,诱导神经炎症反应,促使DA能神经元死亡(Ransohoff,2016),诱导PD。神经炎症又可反向促使α-Syn错误折叠(Gao et al.,2008),加剧其神经毒性效应。α-Syn可抑制NFs的表达,诱导神经元的退行性病变。研究表明,α-Syn的过表达可导致BDNF的表达下调(Yuan et al.,2010)。α-Syn可通过阻断BDNF/TrkB信号通路,诱导DA能神经元的死亡(Kang et al.,2017)。运动干预则可通过促进BDNF的表达进而下调α-Syn,抑制PD。Tuon等(2012)研究发现,8周跑台运动可促进PD小鼠纹状体中的BDNF表达,抑制α-Syn的表达,改善PD病理症状。Zhou等(2017)研究亦发现,3个月自主运动亦可促进PD转基因小鼠脑内BDNF的表达,进而抑制α-Syn的异常表达,改善PD小鼠的认知和运动能力。这说明,运动干预可通过提高BDNF的表达,下调α-Syn的表达,缓解PD病症。
3.5 运动可介导NFs降低氧化应激干预PD
神经系统是氧需求量大和对缺氧非常敏感的组织,也是易受氧化应激损伤的组织。氧化应激可诱导神经元的损伤甚至死亡。周红等(2015)研究发现,PD的发病与DA能神经元的氧化损伤及其抗氧化功能降低有关。NFs具有重要的神经保护功能,尤其是BDNF对各种神经损伤及神经退行性疾病均有神经保护作用(Nagahara et al.,2009)。脑的氧化损伤与BDNF表达密切相关。BDNF可通过增强超氧化物歧化酶(superoxide dismutase,SOD)的活性,提高脑的抗氧化能力(Kapczinski et al.,2008)。提高BDNF水平,也可促进PD患者脑的抗氧化能力(宋彬彬等,2016)。运动干预可通过BDNF信号通路上调实验大鼠杏仁核BDNF表达,促进杏仁核抗氧化酶SOD和过氧化氢酶(catalase,CAT)的活性,降低杏仁核的氧化应激,增强对脑的保护作用(崔建梅等,2016)。此外,IGF-1也具有较强的抗氧化损伤功能。Kokoszko等(2010)研究发现,对体外培养的细胞环境中加入IGF-1,可增强培养细胞抵抗氧化损伤的能力。运动也可通过促进IGF-1的表达,抑制ROS的产生,增进机体的抗氧化能力(Xiao et al.,2013)。Tuon等(2012)的动物实验发现,运动干预可通过促进BDNF的表达,增强PD小鼠脑内抗氧化酶SOD和CAT的表达,缓解神经元的氧化损伤。运动还可通过促进PD小鼠纹状体和血液中的GDNF表达,抑制ROS,缓解模型小鼠PD样症状(姜宁等,2012)。
3.6 运动可介导NFs缓解线粒体功能障碍干预PD
线粒体是机体重要的有氧代谢场所和主要的能量产生中心,也是氧化反应中ROS生成较多的部位,易受氧化应激损伤。神经细胞是对缺氧或有氧代谢供能障碍最敏感的组织,神经元线粒体的氧化应激水平对神经元的存活或功能发挥至关重要。神经元有氧代谢中ROS的产生增多,是神经元线粒体功能障碍的重要诱因,可导致线粒体的能量合成障碍,进而导致神经元异常凋亡,在PD神经退化病变中发挥重要诱导作用(Schapira et al.,2014)。此外,Pukass等(2014)研究发现,DA能神经元中α-Syn聚集增多,可降低线粒体呼吸链复合体I的活性,并诱导ROS增多,导致DA能神经元的死亡。运动可通过介导NFs的表达,发挥对神经元线粒体的保护功能。Peeri等(2015)研究发现,运动可促进BDNF的表达增加,增强线粒体的生物合成,并通过促进线粒体的自噬水平加快清除受损线粒体。运动还可通过BDNF增强线粒体的抗氧化酶(崔建梅等,2016),如SOD和CAT等的表达或活性提高,进而促进对ROS的降解。同时,运动可通过BDNF抑制α-Syn的表达(Tuon et al.,2012),降低对线粒体的损害,缓解线粒体功能障碍(图2)。
3.7 运动可介导NFs缓解神经炎症干预PD
PD病理过程中,DA能神经元丢失,神经胶质细胞激活,释放出细胞因子、趋化因子和ROS,促使神经元产生炎症反应(Qin et al.,2013)。脑内反应性胶质细胞长期激活是PD的病理特征之一,其可加剧神经元的受损程度(Qian et al.,2010)。PD患者脑部尸检显示,神经胶质细胞显著激活,白介素(interleukin,IL),如 IL-1β、IL-2、IL-4、IL-6 以 及肿瘤坏死 因 子- α(tumor necrosis factor α,TNF-α)和血管细胞黏附分子(vascular cell adhesion molecule,VCAM)等炎性细胞因子表达水平显著上调(Reynolds et al.,2008)。运动干预可通过促进NFs的表达,抵抗神经炎症,缓解PD。运动可诱导PD患者血浆BDNF水平的显著增加,VCAM和TNF-α水平的显著降低(Zoladz et al.,2014)。 运 动 还 可 通 过 增 加 NFs(BDNF、GDNF、IGF-1等)和DA的表达,抑制神经炎症反应,促进神经可塑性,延缓PD的发病进程(Monteiro-Junior et al.,2015)。综合3.1~3.7,运动干预NFs调控PD的可能机制如图2所示。
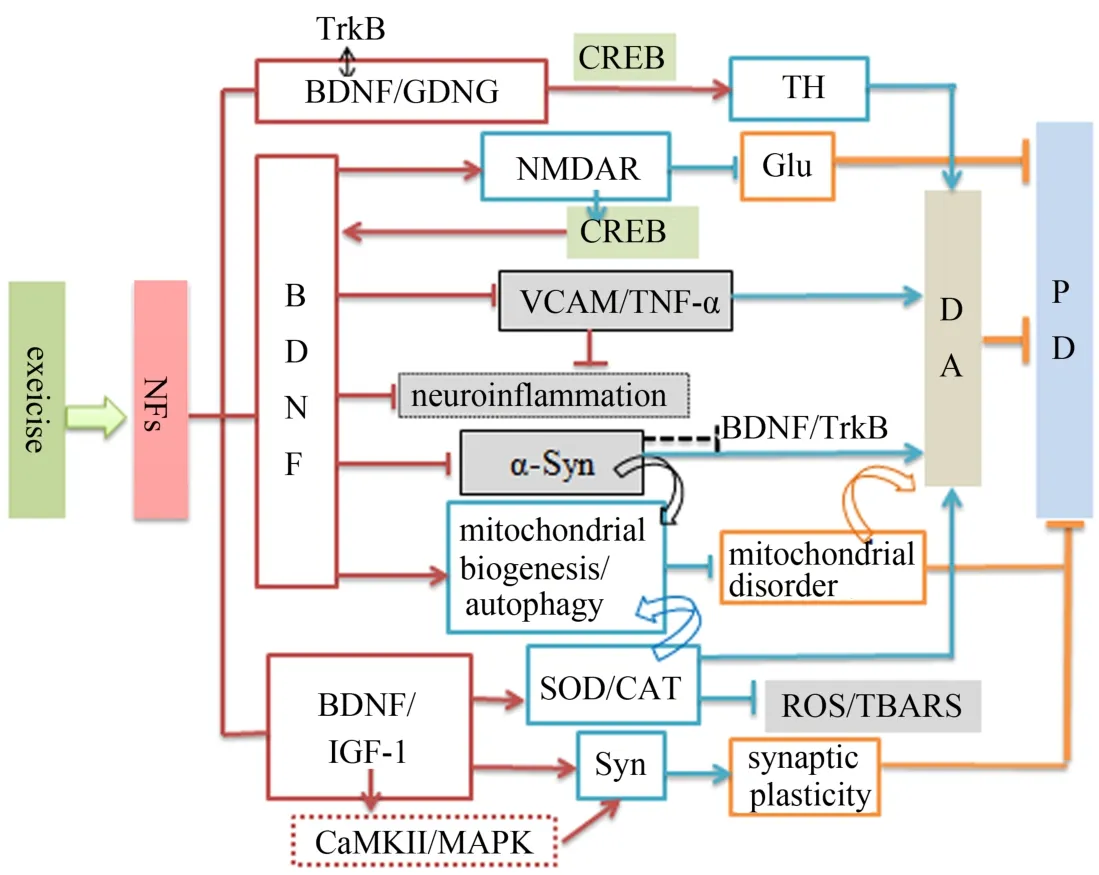
图2 NFs在运动调控PD病理中的可能机制Figure 2.Possible Mechanism of NFs in Exercise Regulating PD
4 不同运动模式介导NFs干预PD的差异分析
运动干预促进神经营养状况的变化对改善PD的病症具有积极作用,但如何确定最适宜的运动量,优化运动干预效果,尚未形成定论。跑台运动是学界公认的PD干预手段。大量动物实验发现,跑台运动可促进NFs的表达,改善实验动物的运动与行为能力。运动干预后,相关NFs,如 BDNF(Fredriksson et al.,2011;Lau et al.,2011;Real et al.,2013;Tajiri et al.,2010;Tuon et al.,2012;Wu et al.,2011)和 GDNF(Faherty et al.,2005;Lau et al.,2011;Speck et al.,2018;Tajiri et al.,2010;Wu et al.,2011)均得到显著性上调。但NFs在运动介导神经保护中的作用也遭到部分研究质疑,这主要与运动强度有关。Schaaf等(2000)研究显示,随着运动强度的增大,机体皮质酮的释放会增加,而皮质酮是BNDF的抑制因子,其可能减弱运动干预的益处。另有研究发现,自主运动较被动运动强度低,对皮质酮的释放影响相对较小(Marlatt et al.,2012;Yanagita et al.,2007)。这说明,自主运动在维持BDNF的表达中效果更佳。陈帅等(2016)通过研究大脑动脉缺血灌注造脑缺血梗死模型大鼠,分别在造模1天、3天和5天后以15 m/min的强度进行跑台训练,发现脑梗死3天和5天后的运动均促进大鼠海马BDNF蛋白表达的显著提高,两组间无显著性差异。但在脑梗死1天后立即进行跑台运动,BDNF表达水平则未见明显增加,说明在脑梗死造模后不同时间段进行运动干预的效果具有差异性。研究还发现,以中等强度(15 m/min)进行训练,可显著提高海马BDNF蛋白水平,而在低强度(10 m/min)或高强度(20 m/min)运动训练后,海马BDNF表达并未显著增加。这表明,BDNF对运动强度反应的敏感性较高,过低或过高强度的运动均不能有效促进BDNF表达,仅中强度训练才能有效增加BDNF的表达。
运动时间也是影响干预效果的重要因素。Wu等(2011)研究发现,对LPS造模的PD小鼠进行1周、2周和4周的跑台运动干预,结果显示,4周的运动干预有效抑制了纹状体中BDNF和GDNF的表达,1周和2周的运动不如4周的运动效果显著。Gerecke等(2010)发现,通过BDNF基因缺失小鼠和野生型小鼠同时注射MPTP造模PD小鼠的实验证实,仅野生型小鼠表现出对MPTP毒性的神经保护作用,BDNF基因缺失的小鼠缺乏抵抗MPTP神经毒性的能力,同时研究分别观察了30天、60天和90天的自主运动干预对注射MPTP的神经保护情况,发现30天的自主运动没有显著的神经保护作用。与30天运动相比,60天的自主运动使DA能神经元的损失降低了16%,而90天的自主运动使DA能神经元仅损失了9%。这表明,长期运动对DA能神经元的保护作用更好,对缓解PD病症的效果更显著。
对PD患者人体试验的运动干预主要包括经典的自行车蹬车运动干预模式和IRT、MRT或MIRT等改进的新型运动康复型干预模式。Zoladz等(2014)和Marusiak等(2015)研究了为期8周的蹬车实验,Sajatovic等(2017)研究了为期12周的蹬车实验,均发现中长期的自行车蹬车运动增加了PD患者血液中的BDNF水平,改善了PD患者的量表测试水平和运动功能。Frazzitta等(2014)采用IRT干预模式、Angelucci等(2016)采用MRT干预模式和Fontanesi等(2016)采用MIRT干预模式的研究均发现,4周的短期新型运动康复干预模式也促进了PD患者血液中的BDNF表达和PD相关量表的测试能力,同样改善了PD患者的运动能力。由此可知,中长期的自行车蹬车运动和短期的新型运动康复干预模式,均可通过促进PD患者血液中的NFs(主要为BDNF)表达,改善PD患者的躯体运动功能,缓解PD相关病理症状。但相关研究尚不深入,还缺乏运动调控其他NFs介导PD病理的研究。
5 结论与展望
PD作为一种常见的神经退行性疾病,患者表现出中脑黑质DA能神经元进行性丢失以及多种NFs的表达不足或功能缺失。NFs是PD病理干预中的重要靶点,BDNF和GDNF是NFs家族中研究最多的成员,与DA合成和DA能神经元的功能维持关系密切。IGF-1和VEGF也是NFs家族的重要成员,分别参与神经元突触发生和脑血管增生等过程。运动能通过调节NFs的表达促进DA能神经元的存活,缓解和改善PD病症。本文综述了BDNF、GDNF、IGF-1和VEGF等几种与PD密切相关的NFs及其在运动调控PD中的可能机制(图3)。运动可调节NFs的表达,作用于神经元,并通过相关的分子信号转导,改善脑的神经可塑性,促进神经血管增生,增强神经元的增殖与存活,缓解氧化应激损伤、线粒体功能障碍和神经炎症,从而实现对PD的调控。
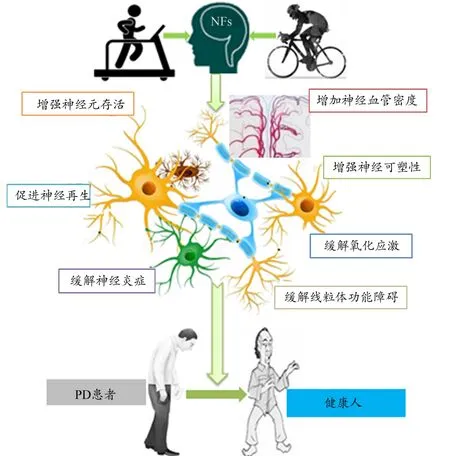
图3 运动介导NFs调控PD的可能机制Figure 3.Possible Mechanism of Exercise Mediated NFs in Regulating PD
在运动介导NFs干预PD的研究中尚存在一定的不足之处:1)生物医学领域的研究已深入PD致病信号通路的探析,而运动干预PD的分子机制还缺乏对相关致病信号通路的深入研究。运动科学应加强与生物医学的学科交叉和融合,发挥不同学科优势,协同促进对PD病理的多维干预。2)PD病理机制复杂,除了中脑黑质和纹状体通路外,还应关注其他神经组织,以便更全面地了解运动对中枢神经系统的神经保护作用。3)DA能神经元是PD脑内的主要受损单位,但神经胶质细胞可能也参与了PD的神经退化过程,但目前运动介导神经胶质细胞调控PD的研究仍未见报道。4)在PD的动物实验研究中,尚不清楚何种运动模式(自主运动或被动运动)与运动强度可对NFs的表达产生更适宜刺激,其他形式运动干预(抗阻运动或水中运动)的实验研究还相对缺乏。因此,应深入探究不同运动模式的差异,综合考虑,设计出更加优化的运动方案,尤其是精确调控运动强度,以避免运动强度失当影响干预效果。
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07 06:21:30
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
自然杂志(2021年6期)2021-12-23 08:24:46
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
汽车维修与保养(2021年8期)2021-02-16 00:28:20
现代装饰(2018年5期)2018-05-26 09:09:01
工业设计(2016年4期)2016-05-04 04:00:15
西南医科大学学报(2015年1期)2015-08-22 13:01:48
电源技术(2015年5期)2015-08-22 11:18:38
