
杜梨砧木留枝嫁接提高鸭梨耐盐能力的机制
2020-06-28 05:54:20孙焕荣李静封晓辉刘小京张秀梅
林业与生态科学 2020年2期
孙焕荣,李静,封晓辉,刘小京,张秀梅
(1 河北省南皮县自然资源和规划局, 河北 南皮 061500;2 中国科学院遗传与发育生物学研究所农业资源研究中心,河北 石家庄 050021)
嫁接通过接穗和砧木细胞的融合、基因交换和特异蛋白合成,可以改变嫁接体的特性,是果树改良的重要手段[1]。嫁接可以提高植株的抗性,如嫁接葡萄可以提高其抗冻和抗病能力,柑橘嫁接可以提高耐盐能力[2]。通过嫁接提高植物耐盐能力的生理机制的研究,主要在果类蔬菜中开展。研究表明,西瓜、茄子和黄瓜嫁接苗对比未嫁接原株具有较高的耐盐能力[3- 4],通过嫁接砧木截留Na+和Cl-等对植物体有害的离子,优化了盐胁迫下植物对离子的吸收和分配,并增加体内渗透调节物质的合成,减少膜质过氧化物的伤害,改变植物激素的合成和运输,从而提高了植物的耐盐能力[1,5]。共生嫁接同样可以提高植株的抗性,如茄子与番茄共生嫁接可以提高植株的抗病能力[6]。但目前为止对果树嫁接提高耐盐能力的生理机制,尚未有报道。
在鸭梨(PyrusbretschneideriRehd.)的砧木中,杜梨(PyrusbetulaefoliaBge.)具有较高的耐盐和耐干旱能力[7-9]。研究表明,以杜梨为砧木可提高梨的耐盐能力[10-12],同时高位嫁接也可以增加梨的耐盐能力[13]。在实际生产中发现,在滨海重盐碱地以杜梨为砧木嫁接梨,可以成活,但是植株盐害程度加重,生长势降低甚至死亡。而杜梨砧木基部偶然生长枝条的植株,却生长良好,有一定的结实。这说明保留砧木枝条可以提高嫁接梨的耐盐能力。但是砧木枝的生长和梨的生长存在竞争,随着砧木枝的增多,梨的生长必然受到限制,因此必须保留合理数量的砧木枝,才能保证最佳的生长结合。由此,为深入探究杜梨留枝嫁接的耐盐机制,验证保留适量的杜梨砧木枝是否将盐分离子吸收储存在杜梨叶片中,并且产生了有利于植物水分利用的物质,以此提高了接合体的耐盐能力等相关问题,本试验通过杜梨砧木留枝嫁接梨,在杜梨砧木上保留不同数量的枝条,比较不同砧木枝处理的生理差异,分析最合适的留枝数量,探讨共生嫁接后的耐盐机制,以期为重盐碱地发展水果产业提供理论参考。
1 材料和方法
1.1 试验地概况
试验地在河北省海兴县中科院滨海农业高效示范区。该地位于渤海湾西岸,由于地势低洼排水不畅,土壤脱盐困难,土壤含盐量4~13 g/kg,年均降水量560 mm,土壤表层盐分含量具有鲜明的季节动态特征,初夏土壤含盐量最高,夏秋季由于雨水淋洗含盐量最低。
1.2 试验材料
本试验将杜梨作为砧木,梨接穗为鸭梨。
试验叶的采集及测量按照叶片类型分为以下3类:(取叶时,每株均按照以下分类采取各类型叶片;测量时,每株中的不同类型叶片重复3次)。
(1)杜梨叶:杜梨砧木枝条上的叶片(包括未嫁接梨的杜梨即杜梨原株和保留不同数量杜梨枝条的已嫁接杜梨的处理)。
(2)功能叶:2011年生长的枝条上的梨叶片,即主要的光合叶。
(3)1 a生枝条叶:取2012年当年生的枝条上的叶片,即新叶。
1.3 试验设计
试验设留砧木枝的个数分别为0、1、2、3,共4个处理,各处理编号分别为T0、T1、T2、T3,每个处理嫁接12株。杜梨留取40~50 cm高的主干截桩,每株嫁接3个接穗,杜梨砧木主干保留不同数量的直径相近的砧木枝。
其中于2010年进行杜梨(当地野生品种)育苗工作,2011年3月移栽,杜梨平均株高100 cm,4月初选择生长一致的杜梨植株嫁接梨。5月调查接穗的成活率,10月测量成活后接穗的长度。
1.4 测定指标及方法
1.4.1 生长量 分别于2011年10月及2012年10月测量植株的接穗长(测定接穗基部到枝条末端的长度),分别用L1、L2表示,二者差值即为枝条生长量,用△L表示。
1.4.2 光合生理 2012年7月选择天晴的时间,在上午9~11时和15~17时用Li-6400以人工光源(光强度为1 200 mol/s),进行光合生理测定。包括光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Cond),胞间CO2浓度(Ci)。同时用SPDA叶绿素含量仪测定叶片叶绿素含量指数;
1.4.3 渗透调节物质含量 采集上述对应的叶片,烘干后用盐酸浸提原子吸收法测定Ca2+,Mg2+,K+,Na+的含量,用AgNO3滴定法测定Cl-含量;用蒽酮和茚三酮比色法,分别测定可溶性糖和脯氨酸的含量。
1.4.4 土壤水、盐含量 用土钻取土分别测定土壤中的水分盐分含量。
1.5 试验地土壤水盐状况
2012年7月测定的不同深度的土壤水分和盐分特征情况,见表1。
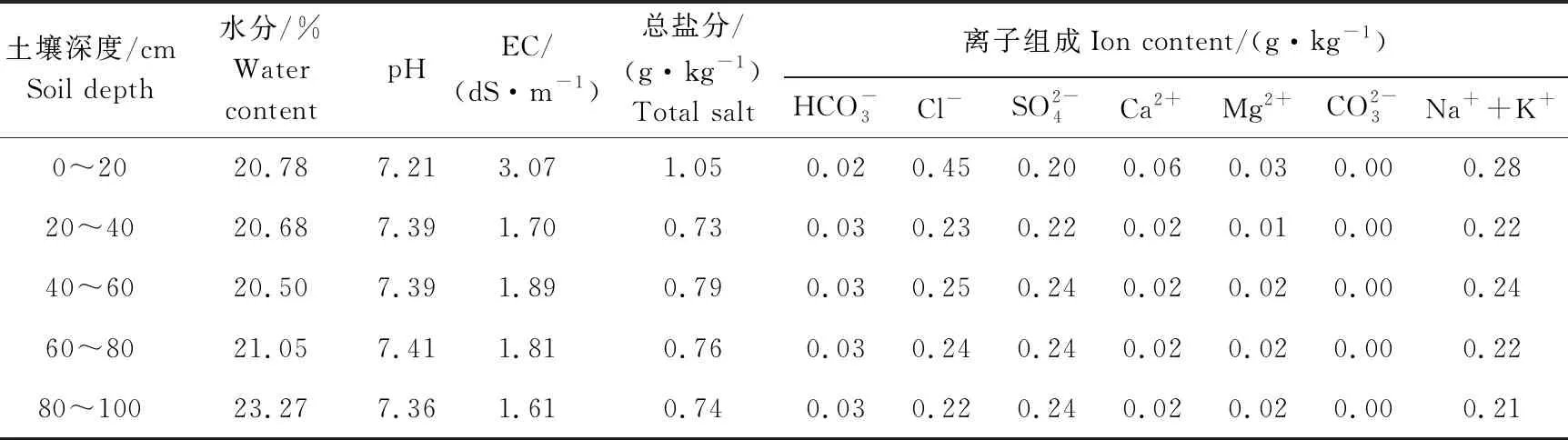
表1 2012年7月土壤水分和盐分特征Table 1 The water and salt content of the soil
1.6 数据整理与分析
采用Excel 2010和Sigmaplot 11.0对试验数据进行整理与统计分析,用Canoco 4.5对叶片中离子含量与光合生理及调节物质进行RDA分析,分析不同类型叶片离子、调节物质和光合生理特征之间的相关性。
2 结果与分析
2.1 砧木枝数量对接穗成活和生长的影响
留取不同砧木数量对接穗成活率和生长量的影响,见表2。
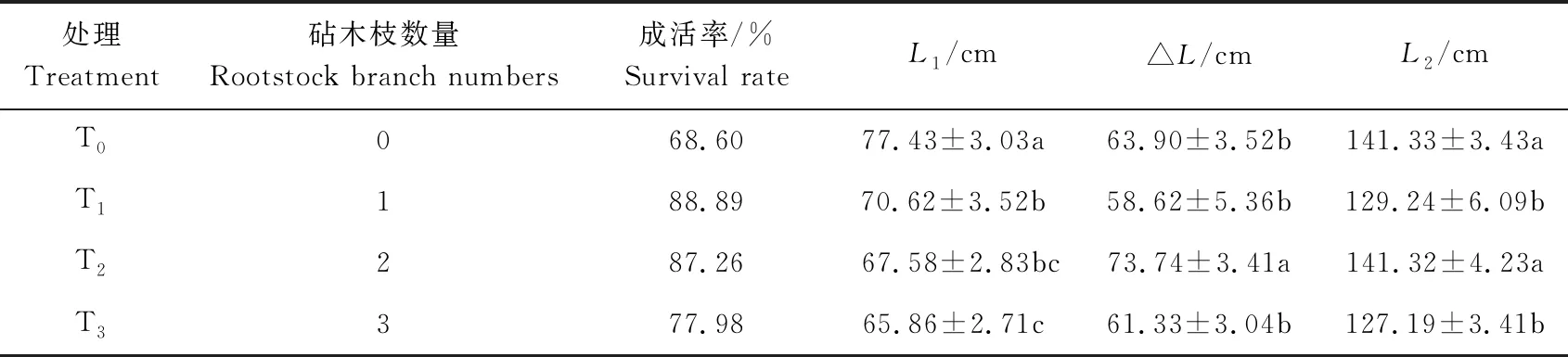
表2 留枝对接穗成活率和生长量的影响Table 2 The effect of reserved rootstock branch on survival rate and the growth of scions
如表2所示,相较于不留枝,保留砧木枝可以提高接穗的成活率,留1枝的处理接穗成活率最高,而不留枝的处理成活率最低,两者相差超过29.57%;但是随着留枝数量的增加接穗的成活率逐渐降低。2011年接穗枝总长L1结果表明,留砧木枝降低接穗的生长量,接穗枝长显著降低,不留砧木枝的接穗生长量最大,而留3枝的生长量最小,留3个砧木枝的处理较不留砧木枝的处理枝长低17.56%。可见砧木枝与接穗之间存在竞争,砧木枝数量越多越不利于接穗的生长。由表中不同处理的生长量△L来看,当保留2个砧木枝时其生长量最大。
2.2 砧木枝数量对叶绿素指数SPAD值的影响
留枝对不同叶片的叶绿素含量(指数SPAD值)的影响不大,如图1所示。不同类型的叶片叶绿素含量存在差异,杜梨叶的叶绿素含量最高,1 a生枝条叶的叶绿素含量最低。但是留取砧木枝数量不同对相同类型叶片的叶绿素含量的影响不显著(图中的杜梨叶中的T0为杜梨原株)。
2.3 砧木枝数量对叶中盐离子含量的影响
留取不同砧木枝数量对叶片中盐离子含量的影响,见图2。

图1 不同留砧数量的叶片中叶绿素指数
Figure 1 The SPAD index of different leaves with different number of branches
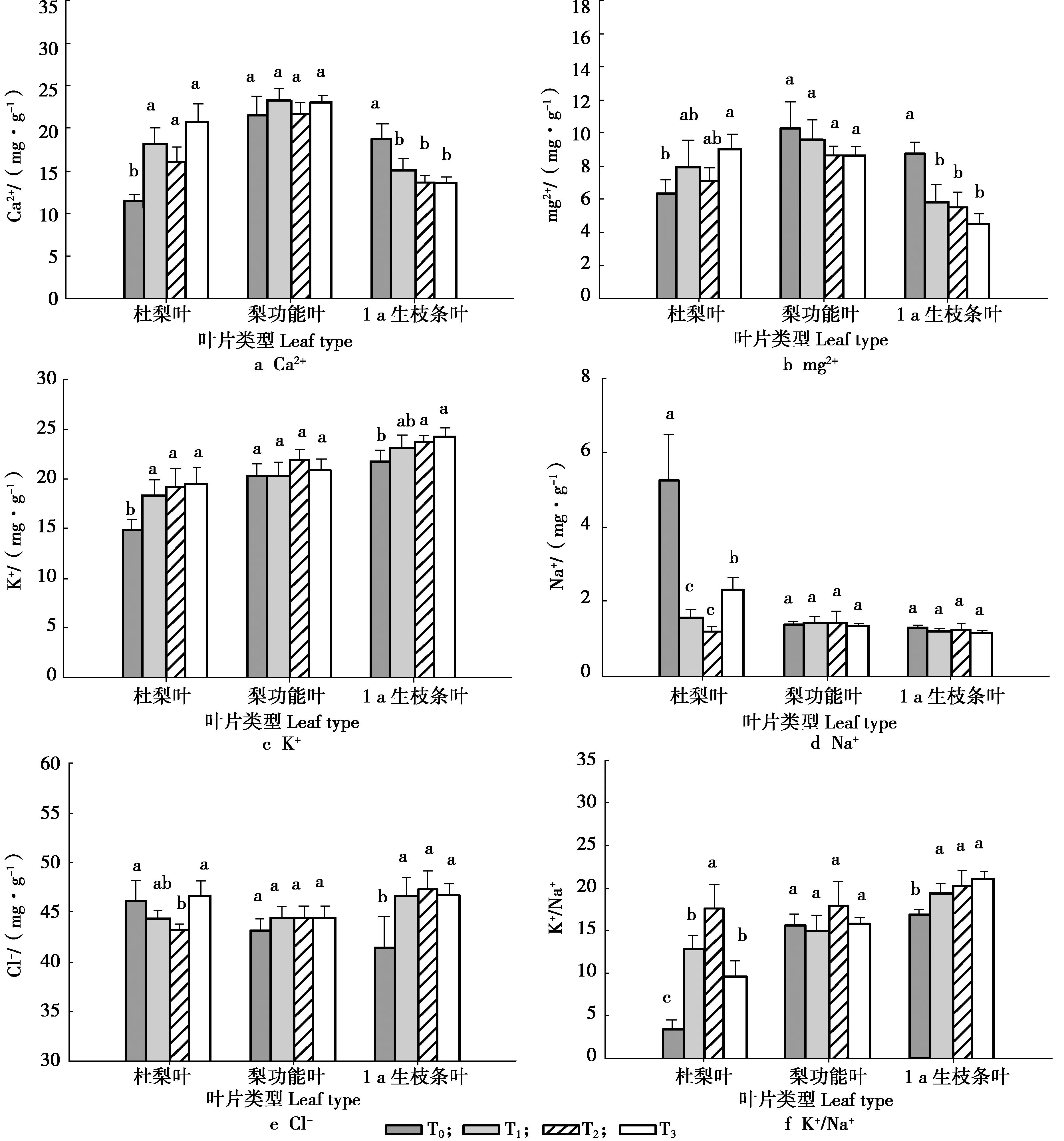
图2 不同留砧数量的叶片中的盐离子含量Figure 2 The salins ions content in leaves with different number of reserved branches
由图2可知,留砧木枝对杜梨叶和1 a生枝条叶的Ca2+含量影响较大。在杜梨叶中,与杜梨原株相比,留砧木枝的叶中Ca2+含量显著较高,但留枝的处理之间Ca2+含量无显著差异;在1 a生枝条叶中,无砧木枝的处理Ca2+含量显著高于留砧木枝的处理,且随着砧木枝数量的增加,叶片中Ca2+含量逐渐下降,无砧木枝的处理较留3个砧木枝的处理高38.77%;留枝数量对功能叶中Ca2+含量影响不显著。不同种类型的叶片中,梨功能叶的Ca2+含量最高。Mg2+含量变化趋势与Ca2+基本一致,功能叶中的含量最高,杜梨叶和1 a生枝条叶Mg2+含量较低。杜梨叶中留砧木枝的叶中Mg2+的含量较杜梨原株高,梨功能叶中Mg2+含量随着砧木枝的增加有逐渐降低的趋势,但是各个处理之间的差异不显著。无砧木枝的处理 1 a生枝条叶Mg2+含量8.34 mg/g,显著高于留砧木枝的处理,比含量最低的3砧木枝的处理高近1倍,且随着砧木枝增加,Mg2+含量呈逐渐下降的趋势。
在不同叶片中,杜梨叶中K+含量最低,1 a生枝条叶中K+含量最高。嫁接梨后,杜梨砧木枝叶中K+显著高于杜梨原株,杜梨叶中K+含量随着砧木枝数量增加而逐渐升高。砧木枝枝数量对梨功能叶中K+含量影响不大,各个处理之间无显著差异;而在1 a生枝条叶中,K+含量随着留枝数量增加而增加,无砧木枝的处理含量最低,只有14.93 mg/g,显著低于有砧木枝的处理。
Na+的含量在杜梨原株叶片中最高,为5.28 mg/g,是留2砧木枝处理的3.46倍,杜梨砧木枝叶中Na+的含量平均只有1.67 mg/g,显著低于杜梨原株。但在梨功能叶和1 a生枝条叶的各个处理之间Na+含量并无显著差异。
同样,不同砧木枝数量的杜梨叶、梨功能叶的Cl-含量差异不显著。但是留砧木枝处理的1 a生枝条叶中Cl-含量显著高于不留砧木枝的处理。
杜梨叶中的K+/Na+差异最大,并且留2砧木枝的处理K+/Na+最高,为17.38,而杜梨原株叶K+/Na+最低,只有3.28。同时留枝显著提高了1 a生枝条叶片的K+/Na+,且随着砧木枝的增加,1 a生枝条K+/Na+逐渐升高;在功能叶中,留2砧木枝处理的K+/Na+最高,但功能叶各处理间K+/Na+的差异不显著。
2.4 砧木枝数量对叶片中渗透调节物质的影响
留取不同砧木枝数量对叶片中可溶性糖和脯氨酸含量的影响,见图3。

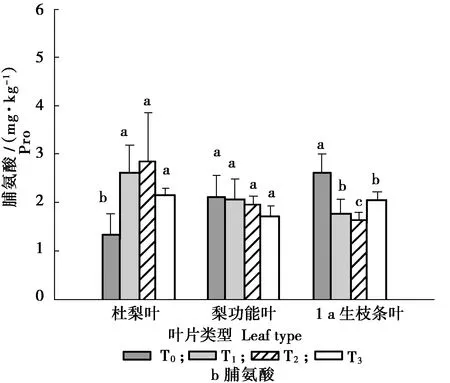
图3 不同留砧数量的叶片中可溶性糖和脯氨酸含量Figure 3 The content of soluble sugar and proline in leaves with different number of reserved branches
由图3可知,嫁接梨后,杜梨砧木枝叶中可溶性糖含量显著低于杜梨原株,且随着砧木枝的增多,杜梨叶中可溶性糖含量逐渐增加;同样,梨功能叶和1 a生枝条叶中的可溶性糖含量都随着砧木枝的数量增加而逐渐增加。说明砧木枝可以提高植株体内的可溶性糖含量。
不同类型的叶片之间,脯氨酸的含量差异较大,杜梨砧木枝叶片中的脯氨酸含量显著高于杜梨原株,其中留2砧木枝处理的脯氨酸含量最高,为3.45 mg/kg,显著高于无砧木枝的处理。而留砧木枝数量对功能叶中的脯氨酸含量无明显影响;无砧木枝处理的1 a生枝条叶中脯氨酸含量最高,为2.62 mg/kg,较留2砧木枝的处理高58.28%。
2.5 砧木枝数量对叶片光合生理的影响
留取不同砧木枝数量对叶片中光合速率、蒸腾速率、胞间CO2浓度、气孔导度含量的影响,见图4。

图4 不同砧木枝数量的叶片光合生理的差异Figure 4 The photosynthetic differnce of leaves with different numbers of reserved branches
由图4可知,不同类型的叶片中,功能叶的光合速率最高,杜梨叶的光合速率最低。杜梨叶的光合生理结果表明,杜梨砧木叶的光合速率显著高于杜梨原株。而在杜梨砧木叶中,光合速率Pn并不随着砧木枝的增加而升高,留2个砧木枝的处理光合速率最高。1 a生枝条叶中,留2个砧木枝的光合速率也显著高于其他处理。随着砧木枝的增加杜梨叶片和功能叶的气孔导度和胞间CO2浓度,都有逐渐增大的趋势。而1 a生枝条叶中,留2个砧木枝处理的气孔导度和胞间CO2浓度最大,光合速率也最大。留2个砧木枝的杜梨叶水分利用效率最高。功能叶中留2砧木枝的处理,光合速率和水分利用效率最高。同时,留枝显著的增加了气孔导度,有利于植物光合,但是留砧木枝的数量越多叶片的蒸腾速率也越高。结果表明,留砧木枝可以显著提高1 a生枝条的水分利用效率,且留2砧木枝的杜梨叶和功能叶水分利用效率也较高。
2.6 叶片中离子含量与光合生理及调节物质之间的关系
在杜梨叶中,光合速率与脯氨酸、Ca2+、Mg2+含量正相关,气孔导度与Cl-和Na+显著负相关。在功能叶中,光合速率与可溶性糖、Mg2+和K+/Na+显著正相关;但是光合速率与气孔导度和脯氨酸的含量关系不大。在1 a生枝条叶中光合速率与K+的含量显著正相关,而与可溶性糖的含量有一定的负相关,与脯氨酸关系不明显(图5)。



图5 不同叶片离子、调节物质和光合生理 特征之间的RDA分析Figure 5 The redundancy analysis between the ions, regulatory substances and photosynthetic characteristics
3 讨论
3.1 保留砧木枝对生长量的影响
在嫁接当年,杜梨砧木枝可以通过主干为接穗提供营养物质,有利于接穗的成活,因此保留砧木枝可以提高接穗的成活率。然而杜梨砧木枝和接穗之间存在对根部养分和水分的竞争,因此砧木枝也抑制接穗的生长,且砧木枝越多,接穗的生长量越小。在嫁接后第2年,砧木枝可以提高当年新生枝条的生长速度,这是由于留砧木枝的植株耐盐能力较不留砧木枝的个体高,留2个砧木枝处理的1 a生枝条的长度,显著高于不留砧木枝的处理,光合特征也印证了留2个砧木枝生长较快。
3.2 留枝对离子分配的影响
杜梨砧木的离子分配特征与杜梨原株有显著不同,未嫁接梨的杜梨叶片中Cl-和Na+较高,特别是Na+含量远远高于嫁接梨叶片中的含量。嫁接后杜梨叶中Cl-和Na+含量则显著下降,而Ca2+、Mg2+、K+的含量显著升高。杜梨叶中的Cl-和Na+含量显著低于杜梨原株,但同时各类型叶片的不同处理的梨叶中的Cl-和Na+含量也无显著差异,说明并不是留砧木枝截留了大量NaCl在杜梨叶中,也没有促进NaCl向上运输到接穗中,因此我们提出的假设被否定,保留杜梨的砧木枝并不能将盐离子储存在杜梨叶片中从而降低对梨接穗中的输送,以降低对梨的离子毒害。而相反,嫁接后杜梨叶片中的Na+和Cl-反而显著降低。
在所有类型的叶中,K+的含量都随着留砧木枝数量的增加逐渐升高,说明砧木枝促进K+的吸收和转运,这有利于接合体抗逆性的提高。研究证明K+在杜梨的渗透调节中有重要作用,K+含量在NaCl胁迫条件下逐渐升高[14]。留2个砧木枝的处理中,杜梨叶和1 a生枝条叶中的K+/Na+较高,当嫁接梨以后,由于梨和杜梨的生理差异,接合体中离子分配发生变化。
3.3 留枝对渗透调节物质和光合生理的影响
脯氨酸、可溶性糖和K+是植物体内重要的渗透调节物质,可降低植物叶片的水势以应对盐分胁迫。而脯氨酸也具有稳定细胞蛋白质结构、防止酶变性失活的作用。随着留砧木枝数量的增加,杜梨和梨叶片中的可溶性糖含量逐渐升高。研究证明盐胁迫下杜梨叶片中的脯氨酸和可溶性糖的含量会升高[15]。在杜梨叶片中可溶性糖含量与光合速率的关系密切,而在梨叶中脯氨酸对光合速率的影响较大,这是由于杜梨叶片中的Na+含量较高,Na+与K+存在竞争,使杜梨叶片中无法积累较多的K+,杜梨叶中积累较多的脯氨酸调节渗透势。而梨叶片中的Na+含量较低,可以积累较多的K+,有利于叶片的光合作用。
3.4 留枝对光合生理的影响
与同属的其他砧木相比较,杜梨具有较高的光饱和点和较高的水分利用效率,嫁接梨后接合体叶片在低盐条件下的光合速率变化不敏感[9]。植物光合作用的限制可分为气孔因素和非气孔因素,前者由于气孔的部分关闭导致的气孔限制,表现为气孔导度下降,导致CO2进入叶片受阻,CO2供应不足,后者因为光合细胞机构和功能受到了损害,而导致叶肉细胞光合活性下降[16]。3种叶片气孔导度与光合速率都为正相关,气孔导度降低使胞间CO2浓度降低,光合作用的底物含量降低,导致光合速率的较低,光合限制为气孔因素。这与高光林的研究结果相同,在低盐胁迫下杜梨与梨的接合体光合作用的限制为气孔因素[10]。
杜梨叶的气孔导度与Cl-和Na+含量显著负相关,与脯氨酸的含量显著正相关,说明在杜梨叶片中Cl-和Na+降低气孔导度,而脯氨酸作为渗透调节物质可以促进气孔的开放,促进光合。而在梨叶片中K+是促进气孔开放的主要物质,脯氨酸的升高反而不利于气孔的开放。杜梨叶片中脯氨酸含量与光合速率正相关,但是1 a生枝条叶中脯氨酸含量和光合速率负相关,说明在杜梨叶中积累脯氨酸有利于光合作用,而在1 a生枝条叶中脯氨酸是在受到胁迫之后产生的,脯氨酸的含量越多表明叶片受到的胁迫越强,光合速率反而降低。留2个砧木枝的处理,功能叶和1 a生枝条叶的K+/Na+较高,K+/Na+也是植物耐盐的重要指标,较高的K+/Na+可提高植物的耐受能力[17],促进了气孔的开放,有利于生长。留2个砧木枝既可以提高耐盐能力,又不因为限制气孔开放而降低光合速率,因此水分的利用效率较高,可以在有限的水分供应中获得较大的生长量。
4 结论
杜梨嫁接梨对砧木叶片的影响较大,杜梨砧木枝与原株相比,生理特征和离子含量发生显著变化。留枝数量对植株的影响是量的变化。留枝虽然对功能叶的影响不大,但可以显著提高1 a生枝条叶的耐盐能力,降低盐害;梨的生长和光合测量的结果都显示:留2个砧木枝既提高了植株的耐盐性又具有较大的生长量,保留2个砧木枝可以均衡砧木和接穗的竞争,既可以提高耐盐能力同时也能实现梨枝条较高的生长量。但是留砧木枝是否对后期梨的生长状况以及果实产量和品质产生影响,有待于继续研究。
猜你喜欢
热带作物学报(2021年9期)2021-11-08 11:55:35
核农学报(2021年3期)2021-02-22 08:47:58
北方果树(2020年6期)2020-11-14 01:35:42
茶叶科学(2018年5期)2018-10-17 01:56:42
作文周刊·小学二年级版(2017年11期)2017-06-21 03:47:16
爱你·阳光少年(2016年10期)2016-05-30 09:06:07
中成药(2016年4期)2016-05-17 06:07:52
现代食品(2016年14期)2016-04-28 08:10:38
食品工程(2015年3期)2015-12-07 10:20:51
西藏科技(2015年10期)2015-09-26 12:10:27
