
阳澄湖原产及“洗澡”中华绒螯蟹背甲的几何形态分析
2020-06-12 12:51薛竣仁陈修报骆仁军刘洪波
水生生物学报 2020年3期
薛竣仁 姜 涛 陈修报 骆仁军 刘洪波 杨 健,
(1. 南京农业大学无锡渔业学院, 无锡 214081; 2. 中国水产科学研究院长江中下游渔业生态环境评价与资源养护重点实验室,中国水产科学研究院淡水渔业研究中心, 无锡 214081)
中华绒螯蟹(Eriocheir sinensis)又称河蟹、大闸蟹, 隶属于节肢动物门(Arthropoda)、甲壳总纲(Crustacea)、十足目(Decapoda)、绒螯蟹属(Eriocheir)[1]。该蟹原产于我国, 目前已入侵至欧洲[2]及美洲[3]。大闸蟹为我国传统的名贵水产品和重要的经济甲壳类动物。2017年其年总产量已经达到了7.5×108kg[4]。其可食部分富含矿质元素[5]、粗蛋白、粗脂肪与氨基酸等[6], 具有较高的营养价值。
我国中华绒螯蟹养殖主要集中在长江流域, 其中江苏省为我国中华绒螯蟹养殖的主产地, 年产量3.5×108kg[4], 约占全国产量的50%[7]; 特别是苏州市阳澄湖中华绒螯蟹因其历史悠久、品优味美最为著名, 且于2005年被原国家质检总局认定为“阳澄湖大闸蟹”地理标志产品实施了原产地保护。值得注意的是, 阳澄湖蟹年产量仅2×106kg, 但市面上销售标称为阳澄湖的蟹量却远超阳澄湖蟹的实际产量[8]。假冒阳澄湖蟹进行不法销售的情况给原产大闸蟹的声誉造成了严重损害。虽然近年来防伪技术不断改进, 但假冒原产蟹的违法行为仍然很难防范[9]; 其中“洗澡蟹”(即将外地蟹移至阳澄湖水域养殖数日, 甚至过水后即冒充“阳澄湖大闸蟹”销售)是最为主要的假冒方式[10,11]。但迄今尚无针对“洗澡蟹”和原产蟹形态及品质异同的比较研究。本研究尝试运用几何形态分析法, 利用阳澄湖实地养殖的原产蟹(从扣蟹一直养到成蟹)及养成于其他湖泊产地, 被笔者特意引入到前者养殖水域“洗澡”的大闸蟹; 对其在相同水域“洗澡”1个月始末时的外部形态进行比较研究, 以期为确认几何形态学特征在区别阳澄湖原产和“洗澡”蟹上的应用潜力提供理论依据和数据支撑。笔者曾对中华绒螯蟹的背甲和腹甲进行过前期的几何形态分析法研究效果的比对。由于对于判别不同产地中华绒螯蟹准确率而言, 腹甲的效果较差(雌、雄形状相异, 前后者平均分别为83.75%、98.75%), 而背甲的效果很好(雌、雄形状相似, 两者均达100%)[12]; 故本研究中只使用背甲来进行上述比较研究。
1 材料与方法
1.1 实验材料
中华绒螯蟹的阳澄湖成蟹样本分别于2018年9月和10月采自阳澄湖围网养殖区(31°26′N, 120°49′E)。非阳澄湖成蟹样本于同年9月采集于江苏省同属长江流域的另一湖泊围网养殖区。所采的非阳澄湖围网养殖样本一部分个体运回实验室后进行“洗澡”前的形态测量, 剩余个体带放至阳澄湖上述原产蟹相同围网养殖区内“洗澡”养殖1个月后, 与10月阳澄湖原产样本蟹同日采集, 进行“洗澡”后的形态测量。外来“洗澡”蟹两类中华绒螯蟹和原产蟹所使用的苗种均来自于江苏南通如东大闸蟹的苗种场(亲本均为长江水系中华绒螯蟹)。“洗澡”始末时两类蟹各选取规格相近的中华绒螯蟹20只(♀、♂各10只)活体进行上述形态学比较测定(表1),所有阳澄湖原产蟹均符合国标GB/T19957-2005[13]所定义的原产蟹(即至少从扣蟹养到成蟹保持同一水域)要求。
1.2 地标点的建立与提取
在设定的高度使用数码相机(Nikon COOLPIX P6000)对非阳澄湖9月的样本、“洗澡”后的10月样本和阳澄湖成蟹样本进行拍摄。地标点的选择需特征明显且易识别, 主要可分为3类, 即以不同组织间交点为特征的Ⅰ型地标点, 以结构中凹陷或凸起点为特征的Ⅱ型地标点和以结构极值点为特征的Ⅲ型地标点[14]。利用tpsdig2软件对拍摄并经过处理的背甲照片建立地标点(图1和表2), 获取地标点的x、y坐标值(2D), 建立地表点的数据文件。

表1 本研究中中华绒螯蟹群体的形态测量信息Tab. 1 Morphometric information of E. sinensis populations in the present study (n=20, Mean±SD)
1.3 平均形与薄板样条分析
地标点的有效性利用tpsSmall软件的最小平方和法则回归分析来检验。用tpsRelw进行普氏叠加等操作, 计算出地标点质心距离, 进而获取平均型(Mean shape)、相对扭曲指数矩阵和数据报告。通过tpsRegr软件的薄板样条分析, 获取成蟹背甲的网格图, 以用于把握背甲的形态变化特征[12]。
1.4 判别分析
采用Bayes法基于各样本的相对扭曲得分(Relative warps scores)实施逐步判别分析。用SPSS 23.0进行相关统计学分析。
2 结果
2.1 几何形态分析
9月和10月中华绒螯蟹背甲地标点数据的切空间距离(Distance in tangent space,y轴)与普氏距离(Procruste distance,x轴)回归系数分别为0.999830和0.999819, 均接近1, 说明本研究所选取的地标点是有效的, 可以应用于后续的几何形态研究。
利用tpsRelw软件在叠印过程中计算出所有样本的平均形(图2)。在对9月中华绒螯蟹背甲地标点数据进行的地标点法分析中, 贡献率最高的地标点分别为11、12、16和17地标点, 共解释了47.80%的形态差异, 其余31个地标点解释了52.20%的形态差异。在对10月中华绒螯蟹进行的地标点法分析中, 贡献率最高的地标点是16、17、18和19地标点, 共解释了42.25%的形态差异, 其余31个地标点解释了57.75%的形态差异。

图1 中华绒螯蟹背甲地标点位置图Fig. 1 Landmark points for morphological measurements on the carapace of E. sinensis

表2 地标点类型与定义Tab. 2 The landmark types and definition
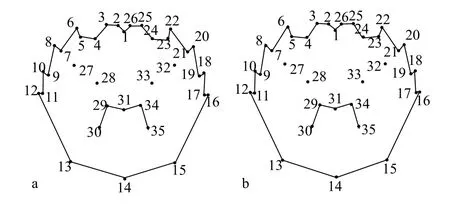
图2 阳澄湖蟹和非阳澄湖蟹背甲的总体平均型Fig. 2 General mean types of the carapace for E. sinensis of Yangcheng Lake and Non-Yangcheng Lake origin
对9月的非阳澄湖和阳澄湖中华绒螯蟹背甲地标点数据进行相对扭曲主成分分析, 前3个主成分的总贡献率为7.72%, 表明各因子线性关系弱, 降维效果差, 无法对总体进行有效的解释。对10月非阳澄湖与阳澄湖中华绒螯蟹背甲地标点数据的相对扭曲主成分分析结果显示, 前3个主成份的总贡献率为7.72%, 降维效果差, 同样无法对总体进行有效的解释。
使用tpsRegr软件将不同产地中华绒螯蟹的差异可视化, 进行网格变形分析后, 从网格图可以发现, 两个产地之间的中华绒螯蟹背甲差异没有非常明显, 所以所有结果均放大10倍进行比较与分析。
从9月份的非阳澄湖蟹与阳澄湖蟹的雄蟹背甲的网格图(图3)中可以发现, 非阳澄湖产中华绒螯蟹的第四侧齿(11、12地标点)向外侧扩展, 背甲后侧缘的3个特征点(13、14、15地标点)向内侧收缩, 阳澄湖蟹的背甲底部更加平缓。非阳澄湖蟹的M型花纹的顶点(29、30、34、35地标点)均向外扩张, 花纹更宽更长, 其余地标点没有显著的差异。
从9月份的非阳澄湖蟹与阳澄湖蟹的雌蟹背甲的网格图(图3)中可以发现, 非阳澄湖蟹额缘的端点(1、2、26地标点)向两边伸展, 顶部更宽。各侧齿之间没有明显的差异, 非阳澄湖蟹后侧缘3个特征点(13、14、15地标点)均向内收缩, M型花纹左下角与右下角的顶点(30、35地标点)均向外伸展,花纹更宽。其余地标点没有显著的差异。
从10月份的“洗澡蟹”与阳澄湖蟹的雄蟹背甲网格图(图4)中可以发现, “洗澡蟹”的第一侧齿(5、6地标点)向内收缩, 第四侧齿(11、12地标点)向外伸展。M型花纹的顶点(29、30、34、35地标点)均向内收缩, 非阳澄湖蟹的M型花纹更短更窄。其余地标点没有显著的差异。

图3 九月中华绒螯蟹背甲网格变形图与变异可视化(变异扩大10倍)Fig. 3 Grid deformation and variation visualization of the carapace for E. sinensis in September (variation enlarged 10 times)
从10月份的“洗澡蟹”与阳澄湖蟹的雌蟹背甲网格图(图4)中可以发现, 阳澄湖蟹的第四侧齿(11、12地标点)均向内收缩, M型花纹(29、30地标点)左侧顶点向外伸展, M型花纹更宽。后侧缘3个特征点(13、14、15地标点)均向外扩展, 底部更加的平缓。其余地标点没有显著的差异。
从图3和图4中华绒螯蟹的差异可视化比较可以看出, “洗澡”前后阳澄湖蟹与本研究所使用非阳澄湖蟹形态差异均主要集中于第四侧齿、M型花纹和后侧缘处。
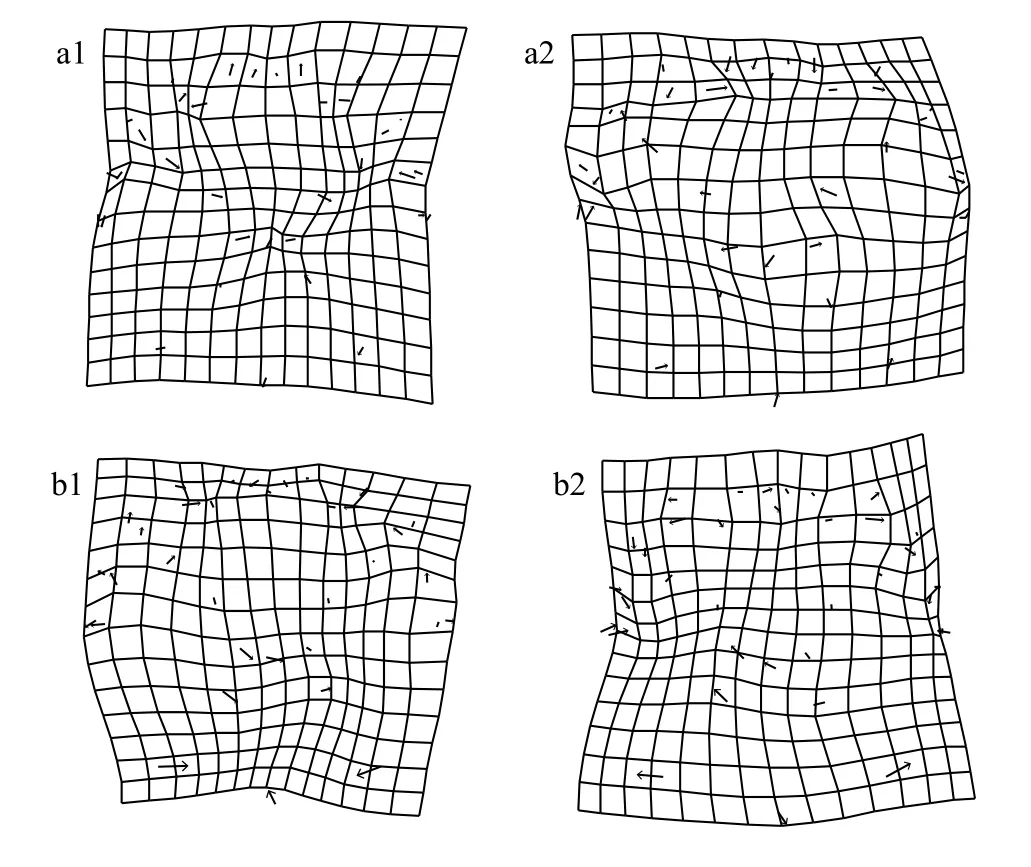
图4 十月中华绒螯蟹背甲网格变形图与变异可视化(变异扩大10倍)Fig. 4 Grid deformation and variation visualization of the carapace for E. sinensis in October (variation enlarged 10 times)
非阳澄湖蟹“洗澡”前后雄蟹与雌蟹的背甲形态均发生了显著变化; 且导致差异的主要地标点趋势相同, 即M型花纹的顶点(29、30、34、35地标点)均向内收缩, M型花纹均变得更短更窄。后侧缘3个特征点(13、14、15地标点)向外侧略微扩展, 底部变得平缓。
2.2 判别分析
通过对9月与10月的阳澄湖蟹与非阳澄湖蟹的逐步判别分析(图5和表3), 可以发现, 对9月与10月阳澄湖蟹与非阳澄湖蟹的判别准确率可以达到100%正确率。对同一产地的雄蟹与雌蟹也可以100%准确的判别。9月与10月的交互验证正确率均可以达到95%。在交互验证中, 9月的非阳澄湖雄蟹有1只误判入阳澄湖雄蟹, 有1只非阳澄湖雌蟹误判入阳澄湖雄蟹。10月的交互验证结果中有两只阳澄湖雄蟹误判入非阳澄湖雄蟹中。
通过对阳澄湖与非阳澄湖“洗澡”前后的雄蟹和雌蟹的判别分析散点图可以发现(图6), 阳澄湖雄蟹在9月与10月已经趋于稳定, 几乎没有发生变化。非阳澄湖中华绒螯蟹雄蟹 “洗澡”前后不仅自身的背甲形态变化较大, 而且与阳澄湖蟹的背甲相比也都存在较大差异。这些形态差异均可以准确的判别出来。对于雌蟹的判别分析散点图中, 阳澄湖蟹变化也较小, 趋于稳定。非阳澄湖雌蟹在“洗澡”后的自身形态变化虽不如雄蟹的变化明显; 但无论是“洗澡”前、还是后, 与阳澄湖原产蟹的差异均较大, 不会趋同。
3 讨论
3.1 “洗澡蟹”形态差异分析
中华绒螯蟹的消费季节一般开始于每年9月,以10—11月为高峰。这段时间也正是“洗澡蟹”最严重的危害期。为不法利益最大化, 只有非阳澄湖产的成蟹才会被用作“洗澡蟹”。本研究中的两类蟹在“洗澡”始末时均达前人报道的成蟹规格[15]。值得注意的是, 因为两类蟹苗种来源相似、种质相同, 理论上其背甲的形态应该相像; 但实际上在“洗澡”开始前两者之间背甲的形态就已经相异(图5和图6)。郑朝臣等[12]在对长江水系的太湖、洪泽湖、高宝湖、长荡湖、阳澄湖以及巴城、兴化“泓膏”养殖水域和长江口崇明自然水域等地产中华绒螯蟹形态上差异的比较研究发现, 即使来源于种质特点相似(长江水系亲本蟹在射阳所育大眼幼体或在南通/崇明所育扣蟹)的苗种, 在经过至少从扣蟹养到商品成蟹近1年的养殖或生长时间后, 上述8个产地蟹的背甲会出现显著的产地差异性变化, 利用几何形态分析法甚至可获100%的产地判别准确率。因此可以想见, 成蟹规格的“洗澡蟹”与阳澄湖原产蟹在经过了从扣蟹养到成蟹长期的养殖期后,其背甲的形态应该也已形成了对两产地水环境相应的适应性变化。Sarà等[16]通过地标点研究尖吻重牙鲷(Diplodus puntazzo)发现, 养殖于不同环境条件(养殖池单养、远岸网箱单养、与金头鲷(Sparus aurata) 养殖池混养)下, 鱼的形态要在8个月后才有明显的适应性变化。张秀霞等[17]通过地标点法对8个野生唐鱼(Tanichthys albonubes)种群的研究发现, 长期相互独立的生境条件和地理隔离使唐鱼的头部形态出现差异。由此可以看出, 鱼、蟹要改变原有身体形态很可能需要长期栖息(如>8个月)于不同产地生境后才会发生。本研究中“洗澡蟹”仅1个月短期转换生境养殖于阳澄湖中, 虽然可见适应性的变化, 但其原有产地而形成的形态特征并未消失,仍可通过几何形态分析其差异与阳澄湖原产蟹进行准确地区分。

图6 基于背甲的中华绒螯蟹判别分析散点图Fig. 6 The discriminate analysis plots of E. sinensis by carapace geometrical morphometry
3.2 地标点的选择及其对背甲形态判别效果的影响
在对9月中华绒螯蟹背甲的分析中, 贡献率最高的地标点分别为11、12、16和17地标点, 共解释了47.80%的形态差异, 其余31个地标点解释了
52.20%的形态差异。在对10月中华绒螯蟹进行的分析中, 贡献率最高的地标点是16、17、18和19地标点, 共解释了42.25%的形态差异, 其余31个地标点解释了57.75%的形态差异。这些贡献率较高的地标点均为各侧齿内外侧顶点, 属于Ⅱ型地标点。结合本研究结果来看, 所选地标点能够解释不同中华绒螯蟹群体形态学上的主要变异信息(判别准确率达100%, 交互验证结果也高于80%)。在相关研究中, 侯刚等[18]使用地标点法对红棘金线鱼(Nemipterus nemurus)、金线鱼(N. virgatus)、深水金线鱼(N. bathybius)和日本金线鱼(N. japonicus)矢耳石进行研究时, 判别准确率可以达到了91.3%, 其中Ⅰ型与Ⅱ型地标点贡献率较高。与之相比, 冯波等[19]使用地标点法与传统形态学方法对不同种群短吻鲾(Leiognathus brevirostris)进行区分的时候, 发现地标点法的区分效果不如传统形态学方法, 通过分析发现其选取的Ⅲ型地标点较多且解释了主要的形态学变异信息。由此可见在利用地标点法进行不同群体形态学差异比较时, 以Ⅰ型和Ⅱ型地标点为优, 同时为了避免造成信息缺失, 少量选择界限较为明显且不易受到主观因素影响的Ⅲ型地标点(如本研究中地标点14)。
3.3 背甲形态判别“洗澡蟹”的潜力
几何形态分析是形态测量学的一种重要方法[20,21]。目前已广泛应用于招潮蟹(Genus uca)[22]、蓝黑鲮(Labeo calbasu)[23]、兔脂鲤(Leporinus cylindriformis)[24]和梭子蟹(Portunus pelagicus)[25]等生物的形态的种内、种间差异性研究或种群判别的研究中。迄今对于不同产地的中华绒螯蟹的判别,已有从形态学[26—28]、分子生物学[29]和生理生化指标[30—32]等不同角度的尝试, 其中以几何形态分析[12]、元素“指纹”[33]和电子舌[34]所获的产地判别准确率较高, 尤其是几何形态分析具有可进行非致死测定且判别准确的优点。在笔者实验室的前期研究中,郑朝臣等[12]通过地标点法对活体中华绒螯蟹的背甲与腹甲进行分析, 准确地判别出了长江水系8个不同湖泊产地的中华绒螯蟹, 其中对雄蟹背甲、雄蟹腹甲、雌蟹背甲、雌蟹腹甲的判别准确率分别达到了100%、98.75%、100%和83.75%。相比于传统的形态测量较低的判别准确率[35], 不仅在判别准确率上有了明显的提升, 同时对于背甲的判别准确率还达到了100%, 平均的判别准确率达到了运用元素“指纹”与同位素的的[33]判别准确率。
本研究运用地标点法对“洗澡蟹”与阳澄湖原产蟹的背甲进行判别。判别结果显示(图5), 在“洗澡”前, 通过对相对扭曲得分进行逐步判别分析, 对非阳澄湖蟹和阳澄湖蟹的判别准确率可以达到100%, 交互验证的准确率依然可以达到95%。在将非阳澄湖的中华绒螯蟹于阳澄湖中进行1个月的“洗澡”之后, 再次通过地标点法与同月阳澄湖产中华绒螯蟹进行甄别, 逐步判别准确率仍然可以达到100%, 并且交互验证准确率依旧可以达到95%, 有效证明了于阳澄湖中“洗澡”的非阳澄湖产的中华绒螯蟹即使“洗澡”1个月时间, 也无法“洗白”变成阳澄湖原产蟹的背甲形态。
本研究通过几何形态学方法对“洗澡蟹”与阳澄湖原产蟹进行了准确地鉴别, 有效地证明了即使1个月“洗澡”也无法有效地改变非阳澄湖产与阳澄湖原产蟹间的形态差异。几何形态学方法的判别准确潜力非常高并且可以进行差异可视化分析, 并且还拥有快速, 方便和非致死的优点。在今后的进一步研究中, 拟尝试将其应用于中华绒螯蟹生长过程中以探索中华绒螯蟹在某固定的环境条件(特别是水质、水环境微量元素含量、水草丰度、流速及水体交换量、养殖密度等)下达到背甲形态稳定所需要的时间, 并且通过形态差异可视化来深入探索中华绒螯蟹生长过程中具体的形态变化, 以便尝试开发可操作推广的基于几何形态分析的背甲识别技术, 为保护阳澄湖原产中华绒螯蟹提供一条创新性的途径。
猜你喜欢
当代水产(2022年6期)2022-06-29
小学生学习指导(低年级)(2021年12期)2021-12-31
南方农业·上旬(2021年7期)2021-09-06
辽宁省博物馆馆刊(2021年0期)2021-07-23
小学生学习指导(低年级)(2020年9期)2020-11-09
海峡姐妹(2019年10期)2019-11-23
幽默大师(2019年5期)2019-05-14
中国自行车(2018年7期)2018-08-14
解放军健康(2017年5期)2017-08-01
WTO经济导刊(2014年7期)2016-03-22
