
核型多角体病毒对玉米草地贪夜蛾的控制作用研究
2020-06-08 09:41张海波王风良陈永明
植物保护 2020年2期
关键词:草地贪夜蛾
张海波 王风良 陈永明

摘要 草地貪夜蛾Spodoptera frugiperda是严重威胁我国玉米生产的入侵害虫。为科学、有效地防控草地贪夜蛾,本文应用浸卵法和浸叶法,在室内研究了几种核型多角体病毒Nucleopolyhedrovirus(NPV)对草地贪夜蛾的毒力,并利用喷雾法进行了田间控制作用研究。结果表明,几种核型多角体病毒浸卵处理10 s对草地贪夜蛾的卵孵化率无明显影响,但苜蓿银纹夜蛾NPV浸卵对草地贪夜蛾卵孵化有一定的延迟作用,24、48 h的孵化率分别为12.50%、69.17%,显著低于对照;核型多角体病毒浸卵对初孵幼虫的毒力较强,棉铃虫NPV、甜菜夜蛾NPV浸卵后初孵幼虫死亡率最高,分别达92.32%、82.49%;草地贪夜蛾3龄幼虫取食浸过核棉铃虫NPV、甜菜夜蛾NPV的玉米叶片72 h后,其死亡率分别为96.39%、92.82%,甘蓝夜蛾NPV对3龄幼虫也具有较强毒杀作用。同时核型多角体病毒对草地贪夜蛾幼虫发育有一定抑制作用,3龄幼虫取食经棉铃虫NPV、甜菜夜蛾NPV处理的叶片72 h平均虫龄分别为3.00龄和3.36龄,显著低于对照。玉米田喷施甜菜夜蛾NPV、甘蓝夜蛾NPV,药后10 d对草地贪夜蛾的防效分别达到86.03%和82.36%。研究表明,甜菜夜蛾NPV、甘蓝夜蛾NPV杀虫剂对玉米草地贪夜蛾具有较好的防控效果,可以作为其有效防控的生物农药。
关键词 草地贪夜蛾; 核型多角体病毒; 浸卵法; 浸叶法; 控制作用
中图分类号: S 476.1 文献标识码: B DOI: 10.16688/j.zwbh.2019647
Abstract Spodoptera frugiperda is a pest that seriously threatens the safety of corn production in China. For the scientific and effective prevention and control of S. frugiperda, the toxicity of several Nucleopolyhedrovirus (NPV) on S. frugiperda was studied in laboratory using egg immersion and leaf dipping methods. Field control effects were carried out using the spray method. The results showed that the immersion with other Nucleopolyhedrovirus for 10 s had no significant effect on the hatching rate of S. frugiperda eggs, but the immersion with AcMNPV delayed the hatching of S. frugiperda eggs, and the hatching rate of 24 h and 48 h were 12.50% and 69.17%, respectively, which were significantly lower than that of the control. The immersion of eggs with NPVs showed high toxicity to the newly hatched larvae, and mortalities caused by the HaNPV and SeNPV were 92.32% and 82.49%, respectively. When the 3rd instar were fed with corn leaves dipped in HaNPV and SeNPV, the mortalities 72 h post treatment were 96.39% and 92.82%, respectively. The MbNPV also had a strong toxicity to the 3rd instar larvae. At the same time, the NPVs inhibited the development of the larvae of S. frugiperda, the average age of the newly hatched larvae was 3.00 instar and 3.36 instar 72 h post treatment with the HaNPV and SeNPV, which were significantly lower than that of the control. The spray of SeNPV and MbNPV in corn field showed high control effect to S. frugiperda, which were 86.03% and 82.36% 10 day post treatment. The results of this study showed that the SeNPV and MbNPV has good control effect against the S. frugiperda, and can be used as a biological pesticide for effective prevention and control of S. frugiperda.
Key words Spodoptera frugiperda; Nucleopolyhedrovirus; egg immersion method; leaf dipping method; field control effect
草地贪夜蛾Spodoptera frugiperda (J.E.Smith)俗称秋黏虫(英文名 fall armyworm),隶属鳞翅目 Lepidoptera夜蛾科Noctuidae灰翅夜蛾属Spodoptera,原产于美洲热带亚热带地区,2017年被评为全球十大植物害虫之一[1]。草地贪夜蛾具有寄主范围广、繁殖能力强、生活周期短、为害程度重、迁飞扩散快、防控难度大等特点[2-3],可为害包括玉米、水稻、甘蔗、高粱、小麦等在内的76科353种植物[4-5]。随着国际贸易活动的日趋频繁,以及自身强大的迁飞扩散能力和多个生物型[6],草地贪夜蛾现已广泛分布于全球100多个国家和地区。目前入侵亚洲的草地贪夜蛾为玉米型,主要取食玉米、高粱等[7]。我国于2019年1月11日在云南省江城县首次发现草地贪夜蛾,其入侵后在低海拔温热地区繁殖为害,并以云南为虫源基地不断向北扩散[8],5月中旬迁飞至长江和江淮流域,6月份进一步北迁至黄淮海玉米产区,扩散路线与我国的玉米种植带高度重合[9]。草地贪夜蛾在我国的扩散速度、为害面积均创造了空前的记录,远远超过我国已经记录的所有其他入侵害虫,是目前威胁我国秋粮生产的头号危险性害虫[10],防控任务迫在眉睫。
生物防治具有持续控制、作用机制多样、对环境友好、不易引起抗性的特点[11],已成为农业害虫绿色防控的重要措施。害虫生物防治常用的介体有天敌昆虫、微生物以及微生物代谢产物等[12]。我国昆虫病原微生物资源较为丰富,针对草地贪夜蛾的真菌[13]、苏云金芽胞杆菌[14]、亚洲玉米螟核型多角体病毒[15]等的研究已取得了一定成果。病毒因作用机理独特、特异性较强、杀虫毒力高、持效时间长、对环境友好、不产生抗性备受关注。自然界中能够感染昆虫的病毒有15个科600多种[16],而能作为生物杀虫剂进行产业化应用的主要是杆状病毒科Baculoviridae病毒,其中核型多角体病毒 Nucleopolyhedrovirus(NPV)主要以鱗翅目昆虫为宿主[17],被称为害虫的“无形杀手”[18]。病毒既可以在种群中横向传染,引发流行病,形成虫瘟,又可以通过纵向传染杀灭蛹和卵,从而有效控制害虫种群及其危害。棉铃虫NPV是我国第一个商品化生产的病毒杀虫剂,其粉剂和悬浮剂都已推广应用于棉铃虫的防治。病毒杀虫剂采用“以毒攻毒”的全新杀虫理念,对农产品质量、农业生态环境十分安全[19],符合国家农业发展要求,是21世纪最具有发展前景的生物农药之一。
我国独特的地理位置、丰富的植物资源、适宜的气候条件等因素促使草地贪夜蛾定殖并将呈现周年常态化发生态势,对草地贪夜蛾应急防控已转为重要害虫的长期防控[12,20]。草地贪夜蛾正处于暴发期,为控制外迁虫源,降低定殖基数,保护粮食安全,化学防治仍是草地贪夜蛾防控的关键技术[21]。而化学农药对生态环境压力不断加大,且入侵我国的草地贪夜蛾在漫长的迁飞过程中,经历了不同国家、地区长时间的化学防治,积累了丰富的抗性[22],加大了防控难度。国内针对草地贪夜蛾防控的天敌、细菌、真菌相继被发现,而关于草地贪夜蛾病毒研究较少,尤其是核型多角体病毒对草地贪夜蛾控制的研究鲜有报道。本文研究了几种夜蛾科核型多角体病毒杀虫剂对草地贪夜蛾室内毒力及田间防效,旨在进一步明确核型多角体病毒杀虫剂对草地贪夜蛾的作用机制,拓展草地贪夜蛾新的生物防治措施,为科学、绿色防控草地贪夜蛾提供参考和依据。
1 材料与方法
1.1 供试材料
供试昆虫:草地贪夜蛾采自江苏省盐城市大丰区玉米田。收集老熟幼虫,用放有新鲜玉米叶的采虫袋带回室内,在培养箱中用玉米叶片饲养至化蛹,待蛹羽化后将成虫放入100 cm×100 cm×90 cm的养虫笼内,喂以10%的蜂蜜水,并放入盆栽玉米苗供其产卵。饲养温度26℃±1℃、相对湿度70%、光周期L∥D=16 h∥8 h,饲养3代以上供试。
供试玉米:品种为‘东单60,辽宁东亚种业有限公司。2019年5月15日播种,室内饲养及杀虫活性测定使用喇叭口期玉米,田间防效试验在玉米大喇叭口期进行。
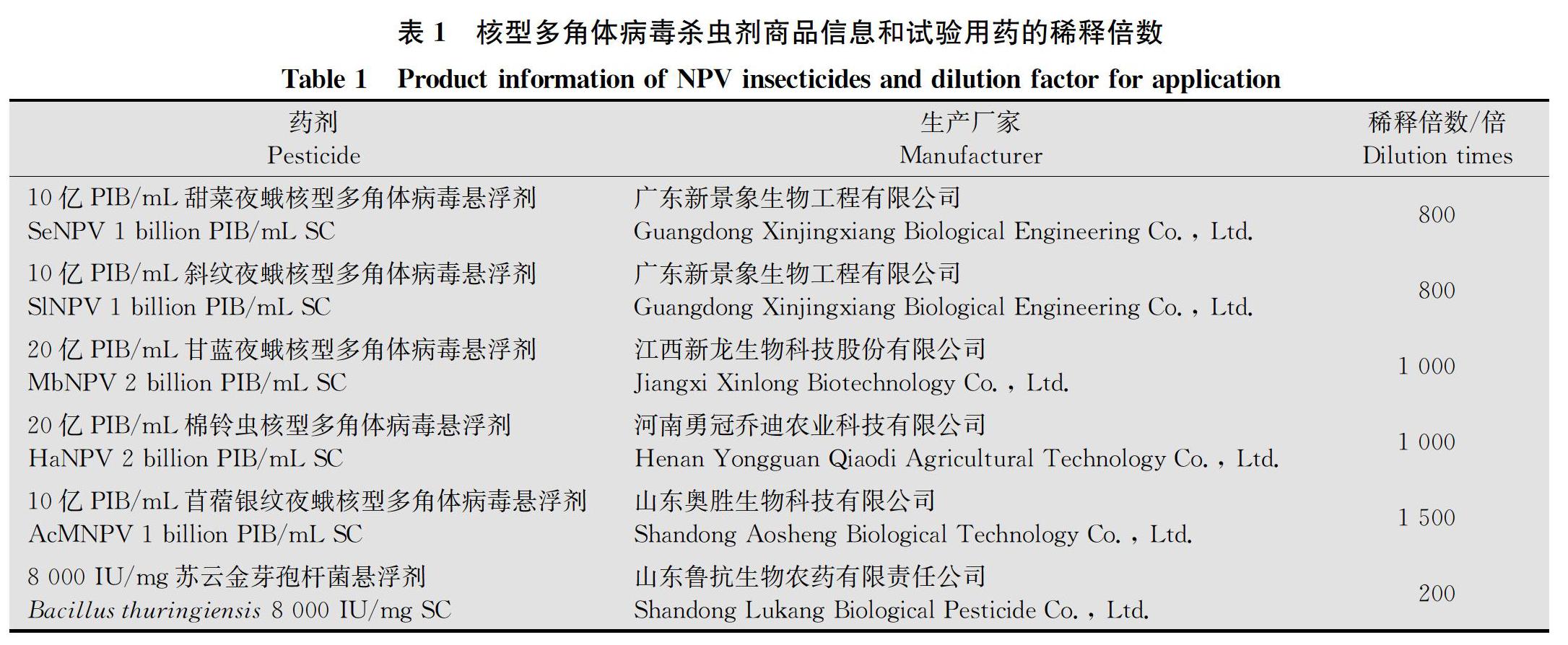
供试药剂:6种供试药剂有效成分、生产厂家及稀释倍数详见表1。稀释倍数以农药登记时的推荐剂量为准,无推荐剂量的农药,以玉米大喇叭口期常规用药量换算。
1.2 试验方法
1.2.1 核型多角体病毒浸卵处理后草地贪夜蛾卵孵化率及初孵幼虫死亡率测定 采用浸卵法。根据陈利民等的[23]方法,略有改动。选取当日产的卵块,用细毛笔将卵块分散为单个卵粒,选取饱满的30粒卵粘于双面胶上制成卵卡,再将卵卡分别在处理药剂中浸10 s,用滤纸吸除多余药剂并晾干后放于直径15 cm的培养皿中,皿中放直径约为5 cm的玉米叶片,以清水处理作对照,各处理重复4次。用体视显微镜每日观察1次,记录卵的孵化情况,计算卵粒孵化率。卵孵化后,每天记录幼虫发育及死亡情况,至幼虫2龄末期止,计算幼虫死亡率。
1.2.2 核型多角体病毒对草地贪夜蛾低龄幼虫的毒力测定 采用浸叶法[24]。将喇叭口期幼嫩玉米叶片剪为长5 cm的叶段,在不同浓度的药液中浸10 s,取出后在室内晾干至表面无游离水。用清水浸渍的叶片作为对照。将晾干的叶片放入直径15 cm的培养皿中,培养皿底部放湿润的滤纸保湿。挑选大小一致、健康的3龄初期幼虫,用镊子接入培养皿,每皿1头,每处理试虫30头,处理重复4次。用保鲜膜封口,并在膜上扎若干小孔透气。放入温度26℃,相对湿度70%,光周期L∥D=16 h∥8 h的光照培养箱中培养。每天定时更换新鲜的浸过药的玉米叶片,于处理后24、48、72 h调查幼虫的存活情况。以尖锐镊子轻触虫体,不能协调运动的个体视为死亡,存活幼虫记录龄期。
1.2.3 核型多角体病毒对草地贪夜蛾玉米田间防效
试验在江苏省盐城市大丰区经济开发区同圣村的玉米田中进行。试验地为砂壤土,pH 7.5,有机质含量1.43%,土壤肥力中等。试验设6种药剂处理,每个处理设4个重复,每个小区面积67 m2,并设清水对照。田间各小区统一常规管理,除试验处理外,其他条件均一致。
按小区实际用药量稀释后采用新加坡利农16 L高压电动喷雾器常规喷雾。施药时先用少量清水充分稀释药剂后,加足相应稀释倍数的水量,进行全株均匀喷雾。田间每个小区随机选择30株受害玉米挂牌标记,不选择小区边缘玉米,调查草地贪夜蛾存活情况,分别于施药前、药后3、7、10 d共4次,记录草地贪夜蛾幼虫活虫数量。施药时间为17:00,施药当天为多云天气。调查期间无明显降雨。
1.3 数据处理
卵孵化率=(孵化卵数/处理总卵数)×100%;
校正死亡率=(处理组虫口死亡率-对照组虫口死亡率)/(1-对照组虫口死亡率)×100%;
校正防效=(处理组虫口减退率-对照组虫口减退率)/(1-对照组虫口减退率)×100%。
试验数据采用DPS软件处理,Duncan氏新复极差法进行多重比较。
2 结果与分析
2.1 核型多角体病毒浸卵处理对草地贪夜蛾卵孵化率及初孵幼虫影响 核型多角体病毒浸卵处理后24 h,草地贪夜蛾卵即开始少量孵化(表2),48 h为卵孵化高峰期,对照组孵化率达82.50%;72 h,除苜蓿银纹夜蛾NPV处理孵化率为89.17%外,其余处理均达90%以上,对照组卵孵化率高达95.83%,各处理组卵孵化率虽均低于对照,但未达到显著水平,说明核型多角体病毒对草地贪夜蛾卵孵化无明显抑制作用;但苜蓿银纹夜蛾NPV浸卵处理后,24、48 h卵孵化率比对照分别低11.67百分点和11.33百分点,均达显著水平(P<0.05),说明其对草地贪夜蛾卵的孵化具有延迟作用。72 h后未孵化的卵大多数干瘪,绝大多数卵不能孵化。
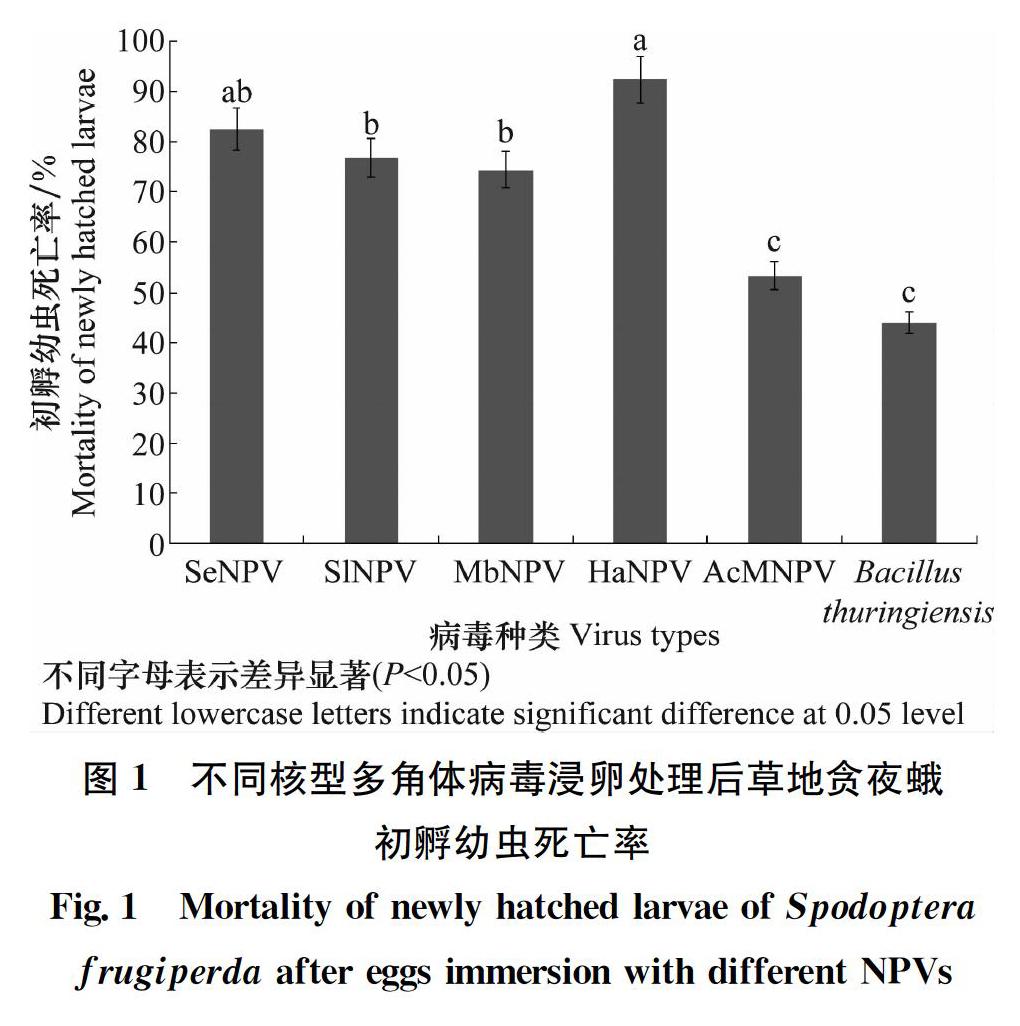
为了进一步明确核型多角体病毒浸卵处理对初孵幼虫的影响,调查了卵孵化后幼虫的死亡率,结果表明,对照组初孵幼虫仅有少数个体死亡,死亡率仅为3.48%,而核型多角体病毒浸卵处理可造成初孵幼虫大量死亡,不同药剂处理的死亡率存在明显的差异(图1)。棉铃虫NPV与甜菜夜蛾NPV浸卵后初孵幼虫死亡率显著高于其他处理(P<0.05),分别为92.32%、82.49%;斜紋夜蛾NPV和甘蓝夜蛾NPV处理后,初孵幼虫的死亡率均达70%以上;苜蓿银纹夜蛾NPV和苏云金芽孢杆菌浸卵处理后,初孵幼虫的死亡率相对较低,分别为53.36%和44.07%。
2.2 核型多角体病毒对草地贪夜蛾低龄幼虫的毒力 不同核型多角体病毒杀虫剂对草地贪夜蛾幼虫的毒力存在明显差异(表3)。草地贪夜蛾3龄幼虫取食经棉铃虫NPV处理的叶片24 h后,死亡率达71.08%,显著高于其他处理(P<0.05),死亡率最低的是苏云金芽孢杆菌和苜蓿银纹夜蛾NPV处理,仅为29.93%和16.10%。
随着取食时间的延长,幼虫的死亡率上升。研究发现,取食72 h,棉铃虫NPV、甜菜夜蛾NPV、斜纹夜蛾NPV对草地贪夜蛾幼虫的校正死亡率分别为96.39%、92.82%、88.32%,显著高于24 h的校正死亡率。苏云金芽孢杆菌和苜蓿银纹夜蛾NPV处理的校正死亡率也分别达到了53.57%、64.10%,较24 h分别提高了23.64、48.00百分点。
取食72 h,各处理存活幼虫的平均虫龄均显著小于对照(P<0.05),其中取食浸过棉铃虫NPV叶片的存活幼虫均停止发育,取食浸过甜菜夜蛾NPV、斜纹夜蛾NPV、甘蓝夜蛾NPV叶片后存活幼虫的发育速率也明显减缓,平均龄期分别为3.36龄、3.41龄、3.58龄。苜蓿银纹夜蛾NPV与苏云金芽孢杆菌对虫龄抑制不明显。
2.3 核型多角体病毒对玉米田草地贪夜蛾的控制作用 田间喷施核型多角体病毒对草地贪夜蛾防效见表4。不同病毒处理后3 d,玉米上草地贪夜蛾的虫量开始不同程度地下降。甜菜夜蛾NPV、棉铃虫NPV处理的校正防效最高,分别为45.20%、42.49%,苜蓿银纹夜蛾NPV处理的校正防效(24.10%)显著低于其他处理,其余处理均达30%以上。可见几种核型多角体病毒杀虫剂及苏云金芽孢杆菌的速效性欠佳。
药后7 d,田间草地贪夜蛾出现死亡高峰,各处理虫口数量均大幅减少,校正防效大幅提升。甜菜夜蛾NPV、甘蓝夜蛾NPV处理校正防效显著高于斜纹夜蛾核型多角体病毒、苜蓿银纹夜蛾核型多角体病毒和苏云金芽孢杆菌(P<0.05),分别为74.99%、70.49%,棉铃虫NPV的校正防效达61.62%,其余药剂处理校正防效也均在50%以上。
药后10 d,各处理对草地贪夜蛾的校正防效仍有不同程度升高。甜菜夜蛾NPV、甘蓝夜蛾NPV处理的校正防效高达86.03%、82.36%,其次是棉铃虫NPV(77.73%),这3种病毒处理的校正防效显著高于其他处理(P<0.05),斜纹夜蛾NPV处理校正防效也在70%以上,苜蓿银纹夜蛾NPV与苏云金芽孢杆菌对草地贪夜蛾防效较差,仅有66.58%、61.19%。
2.4 作物和其他生物安全性
经观察,田间试验过程各处理均未对玉米产生药害和其他不良影响,对有益生物无不良影响。
3 结论与讨论
我国是世界第二大玉米生产和消费国,玉米不仅是口粮资源和饲料资源,更是优良的工业原料。保持玉米的正常供给对促进我国经济健康发展有着极为重要的意义[25]。草地贪夜蛾迁飞入侵我国以来,为害猖獗,对玉米产业将造成重大影响。全国各地研究人员已经在化学防治[26]、生物控制[27]、转基因作物保护[28]、生物种群结构[29]等诸多方面开展了广泛研究,徐磊等研究发现,棉铃虫单粒包埋型核型多角体病毒可诱发草地贪夜蛾细胞株程序性死亡[30]。目前我国尚无登记用于草地贪夜蛾的NPV杀虫剂,因此本文研究了几种夜蛾科已登记的NPV杀虫产品对草地贪夜蛾的控制作用,结果表明,供试的几种核型多角体病毒浸卵处理对草地贪夜蛾卵孵化无明显抑制作用,但对卵孵化均有一定延迟作用,其中苜蓿银纹夜蛾NPV的延迟作用最强。由于初孵幼虫有取食卵壳的习性,因此,NPV浸卵处理对草地贪夜蛾初孵幼虫表现出较强杀伤力,其中棉铃虫NPV、甜菜夜蛾NPV浸卵后初孵幼虫死亡率最高均在80%以上。NPV对草地贪夜蛾3龄幼虫室内毒力测定结果表明,棉铃虫NPV、甜菜夜蛾NPV导致的死亡率在90%以上,甘蓝夜蛾NPV对3龄幼虫也具有较强毒力。同时发现核型多角体病毒对草地贪夜蛾发育也有一定抑制作用。田间防效试验结果表明,甜菜夜蛾NPV药后10 d对草地贪夜蛾防效高达86.03%,农业部推荐的应急用药甘蓝夜蛾NPV防效也相当显著,达82.36%。而常用生物农药苏云金芽孢杆菌对草地贪夜蛾室内毒杀作用及田间防效均表现一般,这与陈利民等[23]、赵胜园等[26]的研究结果一致。
草地贪夜蛾特殊的生物学特性加大了防治难度。田间调查发现草地贪夜蛾在玉米的整个生育期和植株的各个部位均可发生为害。低龄幼虫(1~3龄)取食使玉米叶片形成薄膜“窗孔”,其可在风或其他外力作用下吐丝转移到新的植株上为害,高龄幼虫(4~6 龄)钻入玉米心叶取食使叶片形成不规则孔洞,并排泄大量虫粪,甚至造成整株玉米折断枯死[31]。草地贪夜蛾具有极强的迁飞扩散能力[33],短短几个月已蔓延至全国25 省(区、市)发生面积超过1 00万hm2[32]。一般而言,当外来生物初次被发现时,使用化学防治等应急防控手段短时间内迅速扑杀入侵生物种群被认为是有效的。然而,由于草地贪夜蛾极强的迁飞扩散能力,其入侵速度和为害面积远超人们的预料,仅依靠应急防控措施已难以在短时间内完全扑杀入侵种群。并且草地贪夜蛾在美洲原生地发生已有上百年的历史,化学农药的使用也有几十年的历史[25],其对有机磷类、拟除虫菊酯类、氨基甲酸酯类、抗生素、鱼尼丁受体类农药以及Bt转基因植物均已产生抗性[34],迁飞无疑会导致抗性基因扩散,更加放大了抗药性问题。
根据全球范围内草地贪夜蛾的治理经验,生物农药是一种对草地贪夜蛾有效的控制手段。病毒杀虫剂的杀虫机理与其他生物源杀虫剂有本质的区别,至今没有任何证据表明宿主昆虫对病毒产生抗性[35]。因此,在害虫对常用化学农药产生严重抗药性的地区施用病毒杀虫剂效果会更加突出。核型多角体病毒杀虫剂虽特异性较强,但对近缘的几种靶标害虫也有一定控制作用,甘蓝夜蛾NPV、苜蓿银纹夜蛾NPV是相对广谱的病毒杀虫剂,被用于小菜蛾、甜菜夜蛾、斜纹夜蛾等多种鳞翅目害虫的防治,且效果显著[36]。本研究也发现甜菜夜蛾NPV、棉铃虫NPV、甘蓝夜蛾NPV对草地贪夜蛾有较好的控制作用。昆虫通过取食感染病毒,病毒多角体会在昆虫中肠的碱性环境中溶解释放感染性病毒粒子,并迅速复制导致幼虫死亡[37]。核型多角体病毒作用方式为胃毒,在喷雾时应对玉米的心叶、幼嫩叶的叶面和叶背等害虫喜欢咬食的部位重点喷洒。冯振群等[38]研究表明,在一定范围内,核型多角体病毒使用浓度高低与防治效果呈正相关,为保证田间使用效果,建议施药时使用低容量喷雾施药器械,并可与飞防结合,开发飞防专用病毒剂型,以达到提高使用浓度且不增加药剂成本的目的。
核型多角体病毒杀虫剂虽然是一种新型的生物农药,但在田间应用过程中存在杀虫谱窄、速效性差和效果不稳定等问题。所以应不断扩大病毒筛选范围,获得更多高效、安全的野生型病毒;通过与其他类型农药进行复配拓宽其杀虫谱、延缓化学农药抗性,弥补其速效性差的短板[39]。核型多角体病毒杀虫剂在实际应用过程中受靶标害虫虫龄、温度、紫外线强度、酸碱度等外部因素影响较大[40],这可能是NPV室内和室外对害虫防治效果有所差异的重要原因。所以在使用过程中要注意“打早打小”,龄期越低,防治效果越好,在卵孵化盛期或1、2龄时施药最佳;阳光中的紫外线能使病毒变性失活,故施药时间应在下午4时后,避免阳光直射;碱性环境会溶解NPV的多角体蛋白,因此病毒不得与碱性农药混用,与非碱性农药混用时也应现配现用。科学合理地使用病毒杀虫剂不仅能有效控制害虫的发生与为害,还具有减少化学农药使用、延缓抗性产生、保护农田生态环境、提升农产品质量的优点。随着分子生物学技术和病毒制剂的不断成熟,核型多角体病毒杀虫剂的产业化不断发展,利用核型多角体病毒对我国草地贪夜蛾防控应用前景广阔。
参考文献
[1] STORER N P, BABCOCK J M, SCHLENZ M, et al. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda(Lepidoptera: Noctuidae) in Puerto Rico[J].Journal of Economic Entomology,2010,103(4):1031-1038.
[2] 程东美, 黄江华, 徐汉虹, 等. 50 g/L虱螨脲乳油对草地贪夜蛾的室内活性和田间药效研究[J]. 环境昆虫学报, 2019, 41(5):974-978.
[3] 郭志斌, 蒋睿轩, 唐运林, 等. 重庆地区取食高粱的草地贪夜蛾肠道细菌新分离株的鉴定[J]. 西南大学学报(自然科学版), 2019, 41(9): 9-16.
[4] 赵雪晴, 屈天尧, 李亚红, 等. 2019年云南省草地贪夜蛾春夏季发生特征[J]. 植物保护, 2019, 45(6):84-90.
[5] 吴正伟, 师沛琼, 曾永辉, 等. 3种寄主植物饲养的草地贪夜蛾种群生命表[J]. 植物保护, 2019, 45(6):59-64.
[6] CHAPMAN D, PURSE B V, ROY H E, et al. Global trade networks determine the distribution of invasive non-native species [J].Global Ecology and Biogeography,2017,26(8):907-917.
[7] FUNK D J. “Host forms” and host races: terminological issues in ecological speciation [J]. International Journal of Ecology, 2012: 1-8.
[8] 喜超,姜玉英,木霖,等.草地贪夜蛾在云南的潜在适生区分析及经济损失預测[J].南方农业学报,2019,50(6):1226-1233.
[9] 吴秋琳,姜玉英,吴孔明.草地贪夜蛾缅甸虫源迁入中国的路径分析[J].植物保护,2019,45(2):1-6.
[10] 王磊,陈科伟,陆永跃.我国草地贪夜蛾入侵扩张动态与发生趋势预测[J].环境昆虫学报,2019,41(4):683-694.
[11] 林素坤,刘凯鸿,王瑞飞,等.印楝素对草地贪夜蛾的毒力测定及田间防效[J].华南农业大学学报, 2020,41(1):22-27.
[12] 陈万斌,李玉艳,王孟卿,等.草地贪夜蛾的昆虫病原微生物资源及其应用现状[J].植物保护, 2019, 45(6):1-9.
[13] 张维,彭国雄,夏玉先.昆虫病原真菌防控草地贪夜蛾的现状、问题与展望[J].中国生物防治学报, 2019, 35(5):674-681.
[14] 刘华梅,胡虓,王应龙,等.对草地贪夜蛾高毒力的苏云金杆菌菌株筛选[J].中国生物防治学报,2019,35(5):721-728.
[15] 类承凤,姜干明,彭玲,等.亚洲玉米螟核型多角体病毒分离株鉴定及其对草地贪夜蛾的室内毒力测定[J].中国生物防治学报,2019,35(5):741-746.
[16] SZEWCZYK B, HOYOS-CARVAJAL L, PALUSZEK M, et al. Baculoviruses—re-emerging biopesticides [J]. Biotechnology Advances, 2006, 24: 143-160.
[17] 秦启联,程清泉,张继红,等.昆虫病毒生物杀虫剂产业化及其展望[J].中国生物防治学报,2012,28(2):157-164.
[18] 薛贵收,肖娱玉,朱江,等.昆虫杆状病毒杀虫剂的研究与应用进展[J].江苏农业科学,2009(2):8-11.
[19] 蒋洪,周亮,张珈敏,等.改造病毒基因组增强病毒杀虫剂杀虫效率研究进展[J].植物保护,2008,34(1):5-8.
[20] 唐敏,邝昭琅,李子园,等.叉角厉蝽对草地贪夜蛾幼虫的捕食功能反应[J].环境昆虫学报,2019,41(5):979-985.
[21] 宁素芳,周金成,张柱亭,等.贵州省黔东南地区发现草地贪夜蛾的5种寄生性天敌及其两种重寄生蜂[J].植物保护, 2019, 45(6):39-42.
[22] 郭志斌,蒋睿轩,唐运林,等.重庆地区取食高粱的草地贪夜蛾肠道细菌新分离株的鉴定[J].西南大学学报(自然科学版),2019,41(9):9-16.
[23] 陈利民,黄俊,吴全聪,等.绿色杀虫剂对草地贪夜蛾杀虫活性比较测定[J].环境昆虫学报,2019,41(4):775-781.
[24] HARDKE J T, TEMPLE J H, LEONARD B R, et al. Laboratory toxicity and field efficacy of selected insecticides against fall armyworm(Lepidoptera: Noctuidae) [J]. Florida Entomologist, 2011, 94(2): 272-278.
[25] 马千里,吴吉英子,宋紫霞,等.与美国对比分析草地贪夜蛾在我国的危害发展趋势[J].环境昆虫学报,2019,41(5):929-936.
[26] 赵胜园,孙小旭,张浩文,等.常用化学杀虫剂对草地贪夜蛾防效的室内测定[J].植物保护,2019,45(3):10-14.
[27] 王燕,张红梅,尹艳琼,等.蠋蝽成虫对草地贪夜蛾不同龄期幼虫的捕食能力[J].植物保护,2019,45(5):42-46.
[28] 李国平,姬婷婕,孙小旭,等.入侵云南草地贪夜蛾种群对5种常用Bt蛋白的敏感性评价[J].植物保护,2019,45(3):15-20.
[29] 张磊,柳贝,姜玉英,等.中国不同地区草地贪夜蛾种群生物型分子特征分析[J].植物保护,2019,45(4):20-27.
[30] 徐磊,张菁,苏德明.棉铃虫单粒包埋型核型多角体病毒诱发草地贪夜蛾细胞株Sf的程序性死亡[J].复旦学报(自然科学版),1996(6):675-680.
[31] 郭井菲,何康来,王振营.草地贪夜蛾的生物学特性、发展趋势及防控对策[J].应用昆虫学报,2019,56(3):361-369.
[32] 赵琳超,廖用信,陈壮美,等.不同温度对草地贪夜蛾幼虫和蛹生长发育的影响[J].湖南师范大学自然科学学报, 2020,43(1):41-47.
[33] 杨普云,朱晓明,郭井菲,等.我国草地贪夜蛾的防控对策与建议[J].植物保护,2019,45(4):1-6.
[34] 崔丽,芮昌辉,李永平,等.国外草地贪夜蛾化学防治技术的研究与应用[J].植物保护,2019,45(4):7-13.
[35] 唐璞,王知知,吴琼,等.草地贪夜蛾的天敌资源及其生物防治中的应用[J].应用昆虫学报,2019,56(3):370-381.
[36] 饶楠,梁雪娜,夏红英,等.10亿PIB/mL苜蓿银纹夜蛾核型多角体病毒悬浮剂防治棉铃虫和小菜蛾田间药效试验[J].农药,2018,57(5):377-379.
[37] 孟庆峰,刘晓勇.杆状病毒与昆虫宿主相互作用的研究进展[J].昆虫学报,2013,56(8):925-933.
[38] 冯振群,卢清,李成军.600亿PIB/g棉铃虫核型多角体病毒水分散粒剂防治烟青虫田间试验研究[J].农业灾害研究,2016,6(7):11-12,15.
[39] 邹金城,杨勇,杨益众,等.斜纹夜蛾核型多角体病毒研究进展[J].中国生物防治学报,2016,32(6):800-806.
[40] 單绪南,杨普云.核型多角体病毒的杀虫作用及其田间应用方法[J].中国植保导刊,2009,29(11):39-42.
(责任编辑: 杨明丽)
猜你喜欢
农家科技下旬刊(2019年10期)2019-11-29
植物保护(2019年5期)2019-11-27
植物保护(2019年5期)2019-11-27
现代农业科技(2019年19期)2019-11-21
现代农业科技(2019年19期)2019-11-21
农家科技(2019年10期)2019-11-13
农家科技(2019年7期)2019-10-08
农家科技(2019年7期)2019-10-08
农家科技(2019年8期)2019-09-19
南方农业·下旬(2019年4期)2019-08-26
