
额济纳胡杨空间分布格局及其与微生境异质性的关系研究
2020-06-08 00:38:32高婷婷李景文
林业调查规划 2020年3期
马 季,高婷婷,王 寅,李景文
(1.额济纳旗林业工作站,内蒙古 额济纳 735400;2.北京林业大学林学院,北京 100083)
植物种群空间分布格局的研究是目前生态学的热点问题[1-2]。在不同尺度上,植物的空间分布格局是由多种环境因子长期共同作用的结果,其中生境过滤作用就是森林群落构建的基本驱动力[3]。植物种群的分布格局可分为3类:随机分布、均匀分布和聚集分布[4]。对于不同植物物种空间分布格局已有大量的报道[5-6],但专门针对同一植物种群不同生活史阶段空间分布格局的研究还较少。
胡杨(Populuseuphratica)是中国西北地区唯一能够独自成林的乔木树种[7],由于自然条件的恶化与人为破坏,其生存条件受到严重威胁[8]。胡杨作为荒漠河岸林建群种,是构成内陆河流域河岸林生态系统结构的主体[9]。胡杨的空间分布决定了荒漠河岸林植物群落的结构[10],但目前关于额济纳绿洲胡杨的空间分布格局鲜有文献报道。同时胡杨幼苗天然更新极其困难[11],其种群年龄结构为衰退型[12];对于乔木植物来说,能否在幼苗、幼树阶段成功定植是其存活的关键[13],所以幼苗对环境更为敏感。对不同生活史阶段乔木空间分布格局的成因与过程及其与生境的关系研究,将有利于进一步了解植物种群空间分布格局的形成机制。
本研究通过了解不同生活史阶段胡杨分布格局与微生境因子的关系,分析胡杨在幼苗、幼树、成树3个生活史阶段的分布格局,并对5个环境因子(全氮、全磷、有机质、含水量、全盐)进行空间自相关性分析。以期揭示不同生活史阶段胡杨空间分布格局及其与环境异质性的关系,同时分析影响胡杨分布格局的关键环境因子,阐明微生境异质性在胡杨不同生活史阶段的生态学意义,为进一步掌握额济纳绿洲胡杨的分布状况以及保护问题提供理论基础。
1 研究地概况
研究区位于中国西北干旱区的内蒙古额济纳旗,为温带大陆性气候,年均温为8.9℃,年均降水量为37.9~49.3 mm,蒸发量高达 3 746~4 213 mm,平均蒸发量是降水量的88~109倍。额济纳绿洲核心区的主要土壤类型为非地带性林灌草甸土,并与固定、半固定风沙土、潮土镶嵌分布。额济纳绿洲植被类型是以胡杨和柽柳(Tamarixchinensis)为主的植物群落,其中胡杨林分布面积最大,也是绿洲的重点保护对象[14],胡杨林的生长以及衰退过程与黑河上游来水和地下水埋深状况有着密切联系[15]。
2 研究方法
2.1 样地布设
1984年起开始封育的二道桥林区,覆盖面积约150 hm2,是额济纳绿洲中保护较好的胡杨群落。2015年7月在额济纳胡杨林国家级自然保护区胡杨分布比较集中的二道桥(41°58′7.16″N,101°05′9.96″E)进行样地调查。布设一块100 m×100 m的固定样地,并将固定样地划分为20 m × 20 m的样方(共计25个),采用每木检尺的方法调查所有胸径≥ 1 cm的胡杨个体,记录其胸径、树高、冠幅、相对坐标和生长状况等。
2.2 胡杨不同生活史阶段
根据胡杨种群生活史特点,本文将胡杨种群按胸径大小划分为:幼苗(胸径 < 4 cm)、幼树(4 cm ≤ 胸径 < 10 cm)及成树(胸径 ≥ 10 cm)3个生长阶段[16]。按径级划分的方式分析胡杨的空间分布格局,以判断不同生活史阶段胡杨的分布特征[17]。不同生活史阶段胡杨的分布情况如图1所示。
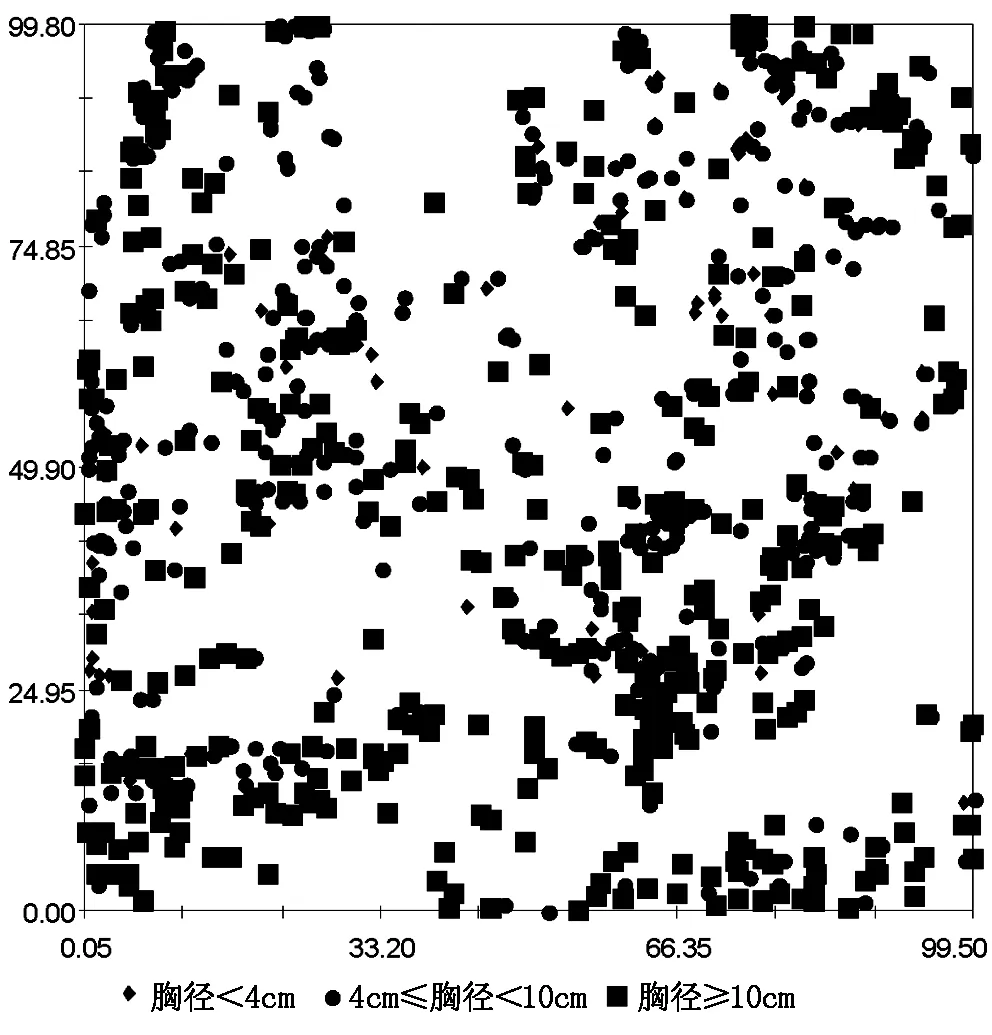
图1 胡杨空间分布
2.3 土壤样品采集与分析
在每个20 m × 20 m的样方中,采用5点取样法分别在样方中央与4个角落采集0~20 cm的土样,记录土壤取样位置的相对坐标。土样剔除植物根系与石砾带回实验室,在室内风干后过2 mm土壤筛,用于测定土壤理化指标。土壤含水率采用烘干法测定;土壤理化性质的测试主要参考鲍士旦[18]研究方法。
2.4 数据分析
运用地统计学中的半方差函数分析法,对胡杨空间分布格局进行定量分析。对部分数据进行对数转换,以保证所有数据符合正态分布。半方差函数分析通过软件 GS+9.0 实现。
采用空间自相关性 Moran′sI指数检验5个微生境因子的空间自相关性。采用类似张春雨等[19]的方法,将样点间最大空间距离120 m设定为有效滞后距离,并划分为8个滞后距离级,最后使用置换检验进行空间自相关指数的显著性检验。空间自相关指数的计算及显著性检验在GS+9.0 及R语言的 spdep 包中完成。
采用冗余分析(RDA)确定不同环境因子对不同生活史阶段胡杨分布格局的作用。最后使用相邻矩阵主坐标(PCNM)分析方法获得空间变量,同时结合环境因子,利用方差分解分析空间变量与环境变量对胡杨分布的相对作用。冗余分析与方差分解使用R软件完成。
3 结果与分析
3.1 胡杨空间分布格局
通过本次调查,样地内共记录胡杨859株,其中幼苗99株,幼树360株,成树400株。研究发现,胡杨成树基台值最大(0.272 6),说明胡杨成树的空间异质性程度较高;成树的半方差函数变程最长(22.4 m),说明胡杨成树具有较强的空间自相关性,且空间自相关强度大于幼树和幼苗;3个生活史阶段半方差函数结构比为0.827、0.869、0.740,说明样地内胡杨分布格局主要受结构性因素影响,由随机因素引起的空间异质性所占比例较小(表1,图2)。
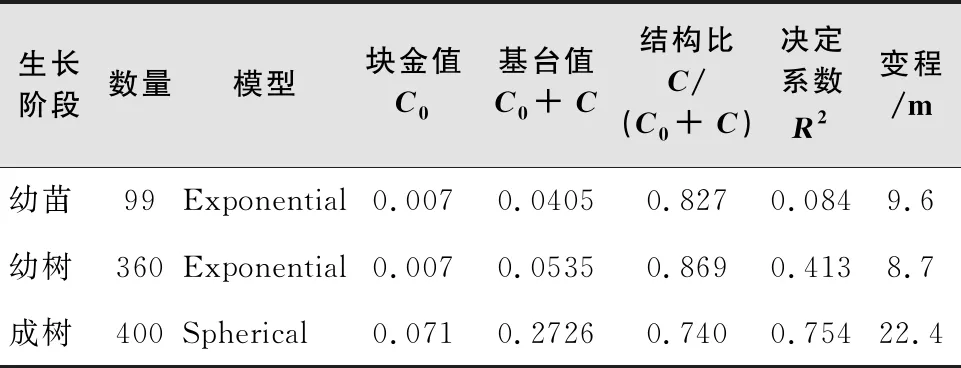
表1 胡杨空间分析的半方差函数模型类型及参数
胡杨幼苗表现出较强的空间自相关结构(图3),当滞后距离级d=1、4、5时,空间自相关指数为正值,表明幼苗分布为空间正相关。同时空间自相关指数随滞后距离的增加不断变化,总体趋势为不断降低,空间正相关逐渐减弱,最终出现空间负相关。而胡杨成树与幼树空间自相关指数则表现为平缓下降,其空间相关性微弱,即空间分布趋于随机分布。
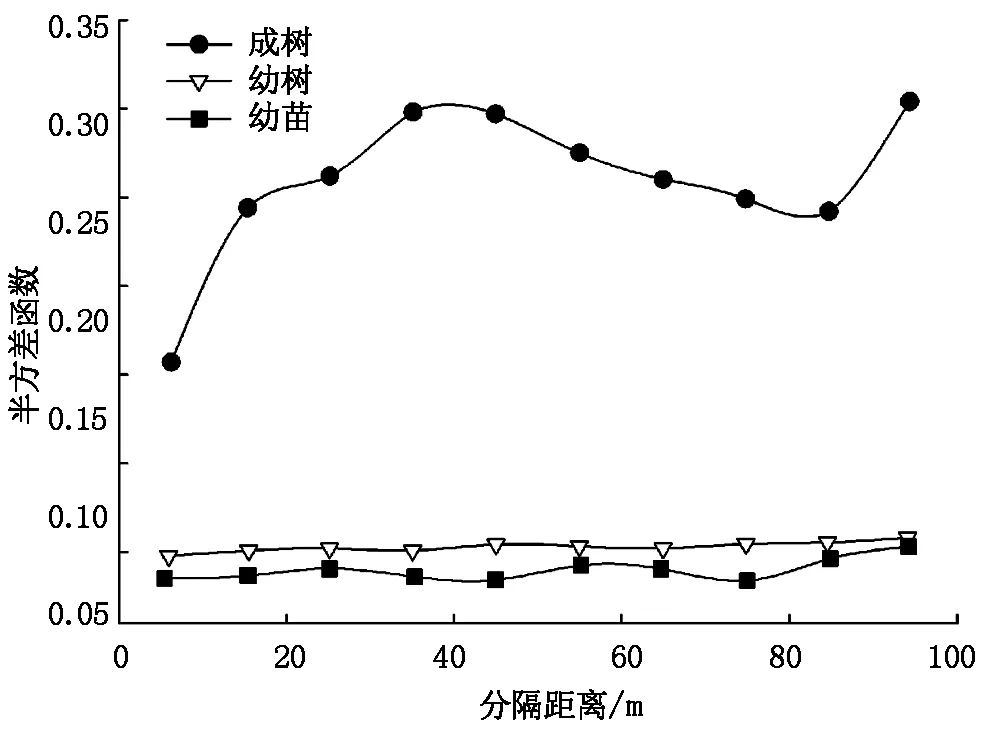
图2 胡杨半方差函数
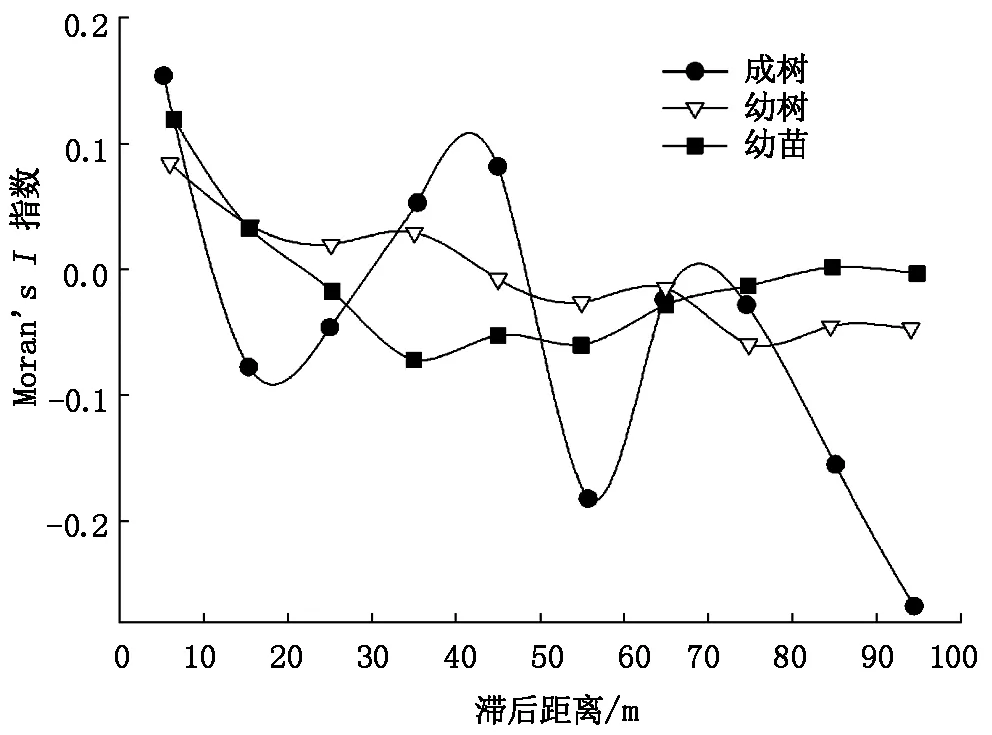
图3 胡杨分布空间自相关指数
3.2 微生境因子的空间分布格局
样地中各土壤理化性质的变异系数均在40%~70%,表现为中等程度变异,其中,土壤全磷的变异系数最大,为70.5%;土壤含水率变异系数最小,为49.6%(表2)。
由表2还能看出,土壤全氮、全磷、含水率最佳拟合模型为指数模型(R2=0.636、0.739、0.913),而土壤有机质、全盐最佳拟合模型为球状模型(R2=0.284、0.852),表明用半方差函数模型能较好地反映样地土壤元素的空间分布格局。各土壤理化指标的块金值(C0)均为正值,土壤有机质、含水率、全盐具有较大的块金值;各指标的块金值与基台值的比值较大(0.503~0.895),说明由结构性因素引起的空间异质性所占比例较高。土壤有机质的半方差函数变程最短(11.4 m),说明其空间连续性较差。

表2 微生境因子半方差函数模型参数
土壤全氮(d=1)、含水率(d=1,4)、全盐(d=1)表现出显著的空间自相关结构,在空间上呈现出显著的聚集分布(表3)。空间自相关指数随滞后距离级的增大逐渐减小;随着尺度的增大,环境变量的分布由聚集趋势转变成分散趋势,空间自相关性从正相关变成负相关。
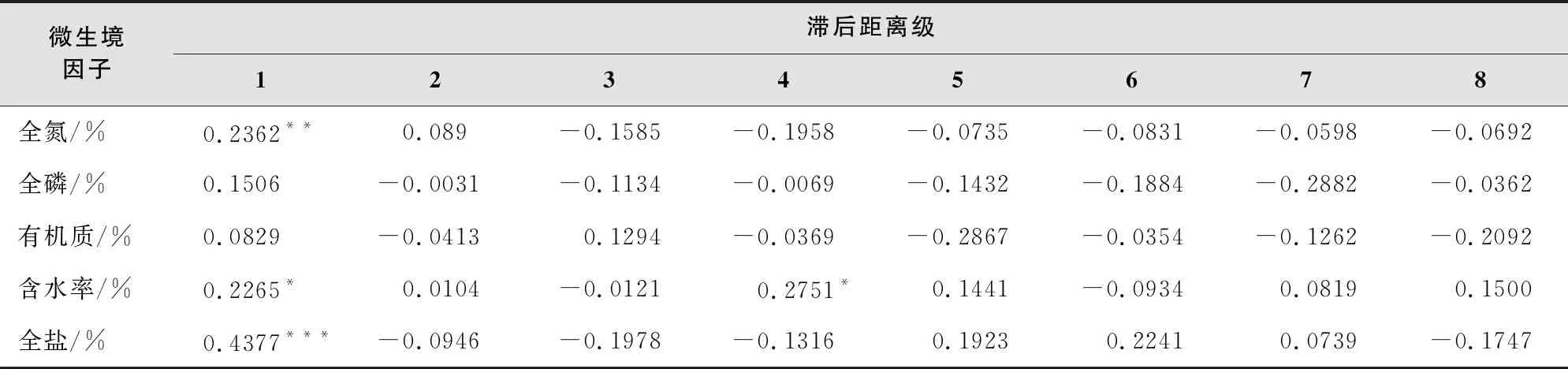
表3 微生境因子的空间自相关分析
注:***代表P<0.001,**代表P<0.01,*代表P<0.05。
3.3 胡杨空间分布格局与微生境因子的关系
5个微生境因子与幼树均无显著关系,而土壤全氮与全盐对成树分布具有显著影响,解释率分别为44.9%、9.2%;土壤全盐与幼苗分布呈负相关关系,其解释率为12.9%(表4)。
利用25个样方的相对位置,运用基于距离的Moran’s特征向量图 (MEM) 构建空间变量,共计获得14个PCNM变量。结合环境变量与空间变量对胡杨的分布进行方差分解(图4)。环境变量对幼苗分布的解释率为12.9%;空间变量对幼树的解释率为36.6%;仅成树受到环境与空间变量的共同作用,且微生境异质性与空间距离的共同作用是成树空间分布格局的主导因素。
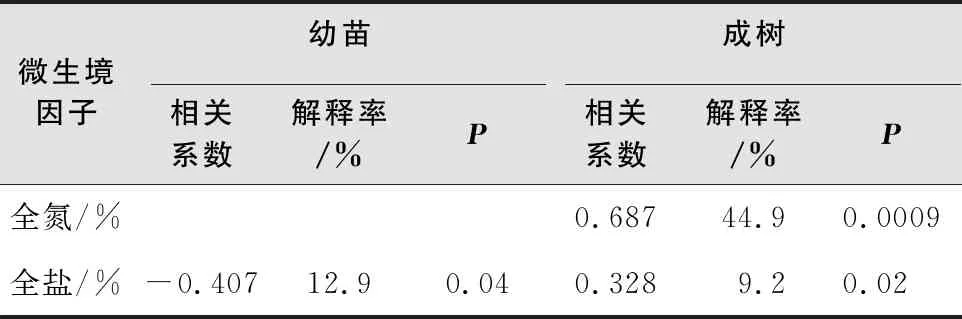
表4 微生境因子与胡杨空间分布相关性及其解释量
注:表中仅显示显著的微生境因子。
4 结论与讨论
种群空间分布是指种群所有个体在水平空间中的分布情况[20]。不同生活史阶段胡杨的空间分布格局存在明显区别,幼苗趋于聚集分布,而幼树、成树则表现为随机分布。胡杨幼苗具有较强的空间自相关结构,而幼树、成树未表现出显著的空间自相关结构,空间异质性主要由随机效应引起。幼苗的聚集分布可能是因为种子扩散的不确定性以及适宜生境较少,仅在少数适宜生境中萌发;同时额济纳胡杨幼苗天然更新极其困难[11],所以胡杨根系主要以根蘖更新为主,根蘖繁殖主要出现在母树周围[21],使得幼苗出现明显的聚集分布[12]。而对于成树与幼树,由于资源限制性作用,种间竞争加剧,个体数量减少,进而增加植物随机分布的可能,目前乔木物种的这种随机分布格局已有大量研究报道[22-23],说明胡杨在不同生活史阶段的分布格局既与额济纳较为特殊且复杂的生境异质性有关,也与胡杨自身的生物学特性和对不同环境的适应策略有关。胡杨空间分布格局与空间自相关因素有关(82.7%、86.9%、74%),其主要受结构性因素的影响,这和李肇晨等[24]的研究结果一致。

图4 微生境变量与空间变量对胡杨空间分布格局的解释
土壤是植物生存生长的基质,对热带雨林树种研究发现,超过30%的乔木树种的分布格局受到土壤养分的强烈影响[25]。本文研究发现,幼苗的空间分布与土壤全盐含量呈负相关,土壤表层盐分含量较高,则会抑制种子萌发与幼苗生长[26]。而成树则与土壤全氮、全盐显著正相关,植物生长必需的养分元素主要来源于土壤中氮、磷,氮素是陆地生态系统植物生长的主要限制因子[27],能够提高植物光合效率[28],胡杨成树的生存与土壤氮含量密切相关。纯粹的微生境变量与空间变量对成树空间分布的解释率较低,而两者共同起到了主导作用;而微生境变量与空间变量对幼苗、幼树的解释率较低,说明一些未知的因素在幼苗、幼树空间分布格局形成过程中起到重要作用。
通过对额济纳二道桥胡杨种群空间分布格局及其微生境异质性的研究,能够直观地反映胡杨种群的分布格局及其与环境的关系。种群的空间分布格局不仅与微生境异质性有关,还受到不同生活史阶段的影响。在今后的研究中,可以通过考虑不同生活史阶段的问题对额济纳胡杨进行后续的研究。
猜你喜欢
金桥(2021年11期)2021-11-20 06:37:30
北广人物(2020年47期)2020-12-09 06:24:12
河北果树(2020年1期)2020-02-09 12:31:18
创新作文(1-2年级)(2018年6期)2018-01-22 12:25:50
现代园艺(2017年11期)2017-06-28 11:32:57
中学历史教学(2016年4期)2016-11-11 07:11:35
甘肃林业(2016年4期)2016-11-07 08:57:01
长江师范学院学报(2016年1期)2016-08-23 11:40:08
中国社会历史评论(2016年2期)2016-06-27 07:11:56
土壤与作物(2015年3期)2015-12-08 00:46:58
