
南滚河自然保护区森林土壤酶活性对海拔升高的响应*
2020-06-02 09:07:08任玉连曹乾斌王志胜
林业科学 2020年4期
任玉连 陆 梅 曹乾斌 李 聪 冯 峻 王志胜
(1.西南林业大学生态与环境学院 昆明 650224;2.云南省林木种苗工作总站 昆明 650215;3.云南南滚河国家级自然保护区沧源管理局 沧源 677499)
土壤酶是森林土壤的重要组分,影响土壤有机物合成、分解与转化等重要生态学过程,对森林生态系统养分循环和能量流动具有重要调控作用(Songetal.,2012;Mayoraetal.,2016)。同时,土壤酶也是森林监测、森林土壤肥力评价以及森林生态系统变化预警的敏感指标(Christensenetal.,1982;王鹏飞等,2017;Badianeetal.,2001)。研究表明,土壤理化性质变化显著影响土壤微生物群落结构(Stricklandetal.,2017)、底物扩散(Koch,1990)和底物的有效性,从而间接影响酶活性(Chen etal.,2017;Wangetal.,2013)。谷晓楠等(2017)研究发现,随海拔梯度变化,植物群落组成、凋落物性质、水热条件、土壤理化性质等均发生相应改变。金裕华等(2011)研究认为,土壤酶活性能敏感地响应不同海拔下土壤碳、氮含量的变化。曹瑞等(2016)也发现,高山峡谷区不同土壤酶对土壤含水量、有机碳、全氮和全磷含量沿海拔梯度变化的响应存在显著差异。海拔梯度变化能够影响降水、温度、土壤养分(Jangetal.,2010;Meganetal.,2016)、植被群落结构(Djukicetal.,2010)和土壤微生物群落组成(Feudisetal.,2017),因此,对沿海拔梯度变化出现的植被类型、微气候和土壤理化性质的变化,土壤酶可能在其结构、组成和活性等方面做出一定响应;同时,调控不同区域土壤酶活性的主导因子可能存在一定差异,使得土壤酶活性对微气候、植物和土壤沿海拔梯度变化的响应存在地区差别。探讨土壤酶活性对植被、气候及海拔梯度变化的响应具有重要意义。
云南南滚河国家级自然保护区地处南亚热带边缘,森林植被群落结构随海拔升高变化明显,且不同海拔植被带的水热条件、植被群落多样性和土壤理化性质存在显著的空间异质性,是研究过渡带森林生态系统结构和功能的理想场地。本研究选取南滚河国家级自然保护区内3种不同海拔梯度典型植被带(沟谷雨林、半常绿季雨林和中山湿性常绿阔叶林),研究植被群落结构、微气候和土壤理化性质沿海拔梯度的变化特征及其与土壤酶活性的关系,进而阐明土壤酶活性对海拔升高的响应特征,以期为理解南滚河国家级自然保护区不同海拔水热条件-植被-土壤理化性质与土壤酶活性之间的相互作用过程和机制提供数据支撑。
1 研究区概况
云南南滚河国家级自然保护区(23°22′57″—23°31′99″E,98°98′87″—99°21′10″N)地处横断山系怒山山脉的南延部分,为滇西纵谷区。气候为南亚热带低纬山地季风气候,夏秋多雨、冬春干旱、干湿季分明。区内地势高低起伏,相对高差2 467 m。年均温差沿海拔升高由22.8 ℃降至9.6 ℃,全年无霜期由365天逐渐降至280天左右,年降水量由1 340.1 mm增至2 592.0 mm。
2 研究方法
2.1 样地设置
在南滚河国家级自然保护区内,选取3种不同海拔梯度的典型植被带,即沟谷雨林(RF)、半常绿季雨林(HF)和中山湿性常绿阔叶林(MF)。每种海拔梯度随机设置3块25 m×25 m重复样地,样地间距约10 m,其基本情况见表1。
2.2 土壤样品采集
2016年5月,运用三点混合取样法采集土壤,分别在每块样地对角线上设置3个1 m×1 m采样点,除去土壤表面凋落物,使用环刀分层采集0~20、20~40和40~60 cm土层土壤样品,剔除石砾和根系等杂物后混合3个取样点的土壤,用四分法取适量土壤装入无菌自封袋,放入4 ℃便携式冰箱保存。将上述土样进行如下处理:一部分样品(200 g)用于测定土壤含水量;一部分(1 000 g)室内自然风干、磨细过筛,用于测定土壤含水量、密度、pH及有机碳、全氮、水解性氮、全磷、有效磷、全钾和速效钾含量;另一部分(1 000 g)冷藏于冰箱中,用于2周内测定土壤脲酶、蔗糖酶、过氧化氢酶、多酚氧化酶和酸性磷酸酶活性。共采集81份土样。
2.3 环境因子测定
气温和降水量数据来源于自然保护区气象站。土壤理化性质测定方法参照《土壤理化分析手册》(中国科学院南京土壤研究所,1987)。土壤含水量采用烘干法测定;土壤密度采用环刀法测定;土壤pH值采用电位法测定(土水比1∶2.5);有机碳含量采用重铬酸钾外加热法测定;全氮含量采用硫酸-高氯酸消化凯氏定氮法(福斯KJ800定氮仪)测定;水解性氮含量采用碱解扩散吸收法测定;全磷和有效磷含量采用钼锑抗比色法测定;全钾含量采用氢氧化钠熔融火焰光度计法测定;速效钾含量采用乙酸铵浸提火焰光度计法测定。每个土壤样品至少重复测定3次,取平均值。土壤温度采用地温计插入不同海拔土层5 cm深处测量,每种海拔土层重复测量3次,取平均值。

表1 样地基本情况
2.4 土壤酶活性测定
土壤酶活性测定分析方法参照文献关松荫(1986)。土壤脲酶活性采用苯酚钠次氯酸钠显色法测定,以24 h内1 g土中NH4+-N的毫克数表示(mg·g-1);蔗糖酶活性采用3,5-二硝基水杨酸显色法测定,以24 h后1 g土中生成葡萄糖的毫克数表示(mg·g-1);过氧化氢酶活性采用KMnO4滴定法测定,以1 g土1 h内消耗0.1 mol·L-1KMnO4的量表示(mL·g-1);多酚氧化酶活性采用邻苯三酚比色法测定,以2 h后1 g土中紫色没食子素的量表示(mg·g-1);酸性磷酸酶活性采用磷酸苯二钠比色法测定,以24 h后1 g土中释放出酚的量表示(mg·g-1)。每个土壤样品至少重复测定3次,取平均值。
2.5 植物群落多样性测定
采用物种多样性指数、均匀度指数和物种丰富度指数测度植物群落的多样性(马克平等,1995)。
Margalef丰富度指数(R)计算方法为:
R=S-1/lnN。
式中:S为种i所在样方不同植被带样地的物种总数;N为种i所在样方采样点各个种的个体数量之和。
Shannon-Wiener 多样性指数(H)计算方法为:
式中:Pi为种i的相对密度,Pi=ni/N;ni为种i的个体数量。
Simpson指数(D)计算方法为:
Pielou均匀度指数(J)计算方法为:
J=H/lnS。
2.6 数据分析
采用Microsoft Excel 2013软件进行试验数据预处理、数据统计和作图;运用单因素方差(ANOVA)分析法检验土壤理化性质和酶活性的差异性,通过LSD法进行显著性多重比较,差异显著性水平设置为α=0.05。利用Canoco 4.5 for Windows软件对土壤酶活性与土壤理化性质间的关系进行分析。由于在对指标进行趋势对应分析时发现梯度长度最大值小于3,因此本研究采用线性模型中的冗余分析进行趋势分析。采用蒙特卡洛检验对土壤理化性质进行重要性排序。对土壤酶活性有极显著影响且解释量在50%以上的12个环境因子进行深入分析,进一步确定单一环境因子对土壤酶活性的影响,分析采用Canoco 4.5中的T-value双序图进行检验,明确各环境因子与各土壤酶活性的关系,若某个土壤酶活性指标的箭头连线完全落入线圈中,则代表该土壤酶活性与土壤理化指标具有显著相关性,其中落入实线圈表示呈显著正相关,落入黑色虚线圈表示呈显著负相关(解雪峰等,2018)。
3 结果与分析
3.1 不同海拔梯度植被带的水热条件及植被群落多样性差异
由表2可知,各植被带的年均气温、年均降水量和土壤温度差异显著(P<0.05)。年均气温和土壤温度随海拔升高而降低,即沟谷雨林>半常绿季雨林>中山湿性常绿阔叶林,而年均降水量随海拔升高而增加,即沟谷雨林<半常绿季雨林<中山湿性常绿阔叶林。

表2 不同植被带水热条件特征①
① 同列不同小写字母表示差异显著(P<0.05),最大的平均数标记为a。下同。Different lower-case letters in the same column indicates significant differences(P<0.05), the maximun mean value mark as a.The same below.
由表3可知:Margalef 丰富度指数、Shannon-Wiener 多样性指数、Simpson 指数和Pielou 均匀度指数随海拔升高的变化趋势显著不同(P<0.05)。Margalef 丰富度指数、Shannon-Wiener 多样性指数和Pielou 均匀度指数均随海拔升高而减少,即沟谷雨林>半常绿季雨林>中山湿性常绿阔叶林,沟谷雨林与半常绿季雨林和中山湿性常绿阔叶林间差异显著(P<0.05),半常绿季雨林与中山湿性常绿阔叶林间差异不显著(P>0.05);Simpson 指数随海拔升高而增加,即沟谷雨林<半常绿季雨林<中山湿性常绿阔叶林,3者之间差异均显著(P<0.05)。

表3 不同海拔梯度植被带的多样性指数
3.2 不同海拔梯度植被带的土壤理化性质差异
土壤理化性质随海拔升高呈现不同变化规律(表4),且差异显著(P<0.05)。土壤密度(沟谷雨林、半常绿季雨林和中山湿性常绿阔叶林分别为1.38、1.11和1.06 g·cm-3)和pH(沟谷雨林、半常绿季雨林和中山湿性常绿阔叶林分别为5.53、5.13和4.22)随海拔升高而降低;土壤含水量及有机碳、全氮、全磷、全钾、水解性氮、有效磷和速效钾含量则随海拔升高而增加,中山湿性常绿阔叶林的土壤含水量及有机碳、全氮、全磷、全钾、水解性氮、有效磷和速效钾含量分别是沟谷雨林的1.43、6.55、5.31、3.28、3.01、3.7和3.11倍。
不同海拔梯度植被带的土壤理化性质沿土层深度呈现不同变化规律,且差异显著(P<0.05)。半常绿季雨林和中山湿性常绿阔叶林的土壤密度、pH值和C/N均在40~60 cm土层最大,表现为随土层加深而增加;土壤有机碳、全氮、全磷、水解性氮、有效磷含量和土壤含水量在0~20 cm土层最高,在40~60 cm土层最低,表现为随土层加深而减少;沟谷雨林的土壤含水量在0~20 cm土层最高,在20~40 cm土层最低,且土壤C/N在20~40 cm土层最高,土壤密度和pH值均在40~60 cm土层最大。
3.3 不同海拔梯度植被带的土壤酶活性差异
土壤脲酶、蔗糖酶和酸性磷酸酶活性在不同海拔梯度植被带间均差异显著(P<0.05),表现为中山湿性常绿阔叶林(土壤脲酶、蔗糖酶和酸性磷酸酶活性分别为301.13、24.76和7.36 mg·g-1)>半常绿季雨林(土壤脲酶、蔗糖酶和酸性磷酸酶活性分别为168.17、14.25和2.03 mg·g-1)>沟谷雨林(土壤脲酶、蔗糖酶和酸性磷酸酶活性分别为122.03、10.86和0.88 mg·g-1)。土壤脲酶、蔗糖酶和酸性磷酸酶活性均表现为0~20 cm土层最高,40~60 cm土层最低。土壤脲酶活性和蔗糖酶活性在各土层均差异显著(P<0.05);土壤酸性磷酸酶活性除在半常绿季雨林中表现为0~20 cm土层显著(P<0.05)大于20~40和40~60 cm土层外,在其他海拔梯度植被带的各土层间差异不显著。相同土层的土壤脲酶、蔗糖酶和酸性磷酸酶活性随海拔升高呈增加趋势,但也表现出一定差异性。不同梯度土壤脲酶和酸性磷酸酶活性在相同土层差异显著(P<0.05),而沟谷雨林与半常绿季雨林之间的蔗糖酶活性在20~40 cm和40~60 cm土层差异不显著。土壤脲酶、蔗糖酶和酸性磷酸酶活性均随土层加深逐渐降低(图1)。
不同海拔梯度植被带土壤多酚氧化酶活性总体上随海拔升高而减弱,即沟谷雨林(1.18 mg·g-1)>半常绿季雨林(0.62 mg·g-1)>中山湿性常绿阔叶林(0.40 mg·g-1),且随土层加深而降低。除半常绿季雨林多酚氧化酶活性在20~40 和40~60 cm土层间差异不显著外,其他海拔梯度植被带各土层间差异均显著(P<0.05)。不同海拔相同土层的多酚氧化酶活性均以沟谷雨林土壤最高,除沟谷雨林与半常绿季雨林40~60 cm土层间多酚氧化酶活性差异不显著外,其余均有显著差异(P<0.05)(图1)。
土壤过氧化氢酶活性表现为中山湿性常绿阔叶林(1.44 mL·g-1)>半常绿季雨林(0.93 mL·g-1)>沟谷雨林(0.79 mL·g-1),且随土层加深而增加,在40~60 cm土层达到峰值。同一海拔植被带,3个土层间的土壤过氧化氢酶活性差异显著(P<0.05)。不同海拔相同土层过氧化氢酶活性均以中山湿性常绿阔叶林土壤最高,且在0~20和40~60 cm土层沟谷雨林与半常绿季雨林间差异不显著,二者与中山湿性常绿阔叶林间有显著差异(P<0.05);半常绿季雨林与中山湿性常绿阔叶林间20~40 cm土层差异不显著,二者与沟谷雨林间有显著差异(P<0.05)(图1)。
3.4 土壤酶活性与环境因子的相关关系
3.4.1 土壤酶活性与环境因子的RDA排序 表5为环境因子对5种土壤酶活性差异性的解释量,5种土壤酶活性在第Ⅰ和Ⅱ轴的解释量分别为98.8%和0.6%,即前2轴18种环境因子可累计解释土壤酶活性特征的99.4%,且对土壤酶活性与环境因子关系的累计解释量达到100%。这表明前2轴能反映土壤酶活性与环境因子关系的绝大部分信息,且主要由第Ⅰ轴决定。
从土壤酶活性与环境因子的二维排序(图2)可知,土壤全磷含量、含水量、pH值、有机碳含量、年均气温、土壤温度、水解性氮含量、年均降水量、全氮含量和全钾含量的箭头连线较长,能较好地解释土壤酶活性的差异。含水量和全磷含量与脲酶活性、有机碳含量与酸性磷酸酶活性、有效磷含量、全氮含量和水解性氮活性与蔗糖酶活性的夹角小,且呈显著正相关,说明含水量、全磷含量是影响脲酶活性的主导因子,有机碳含量主要影响酸性磷酸酶活性,有效磷、全氮和水解性氮含量主要影响蔗糖酶活性。土壤密度和pH值与脲酶、酸性磷酸酶和蔗糖酶活性呈较强负相关;年均气温、土壤温度、Margalef 丰富度指数、Shannon-Wiener 多样性指数和Pielou 均匀度指数与过氧化氢酶、脲酶和酸性磷酸酶活性呈显著负相关,与多酚氧化酶活性呈正相关;年均降水量与过氧化氢酶、脲酶、酸性磷酸酶和蔗糖酶活性呈正相关,与多酚氧化酶活性呈负相关。

表4 不同海拔梯度植被带的土壤理化性质①
①在同一土层,不同海拔梯度植被带间不同小写字母表示差异显著(P<0.05);在不同土层,同一海拔植被带间不同大写字母表示差异显著(P<0.05)Different lower-case letters indicates a significant differences(P<0.05)among vegetation zones at different elevation gradients in the same soil layer, different capital letters indicates a significant differences(P<0.05)among the same elevation vegetation zones in different soil layers.

图1 不同海拔梯度植被带各土层土壤酶活性
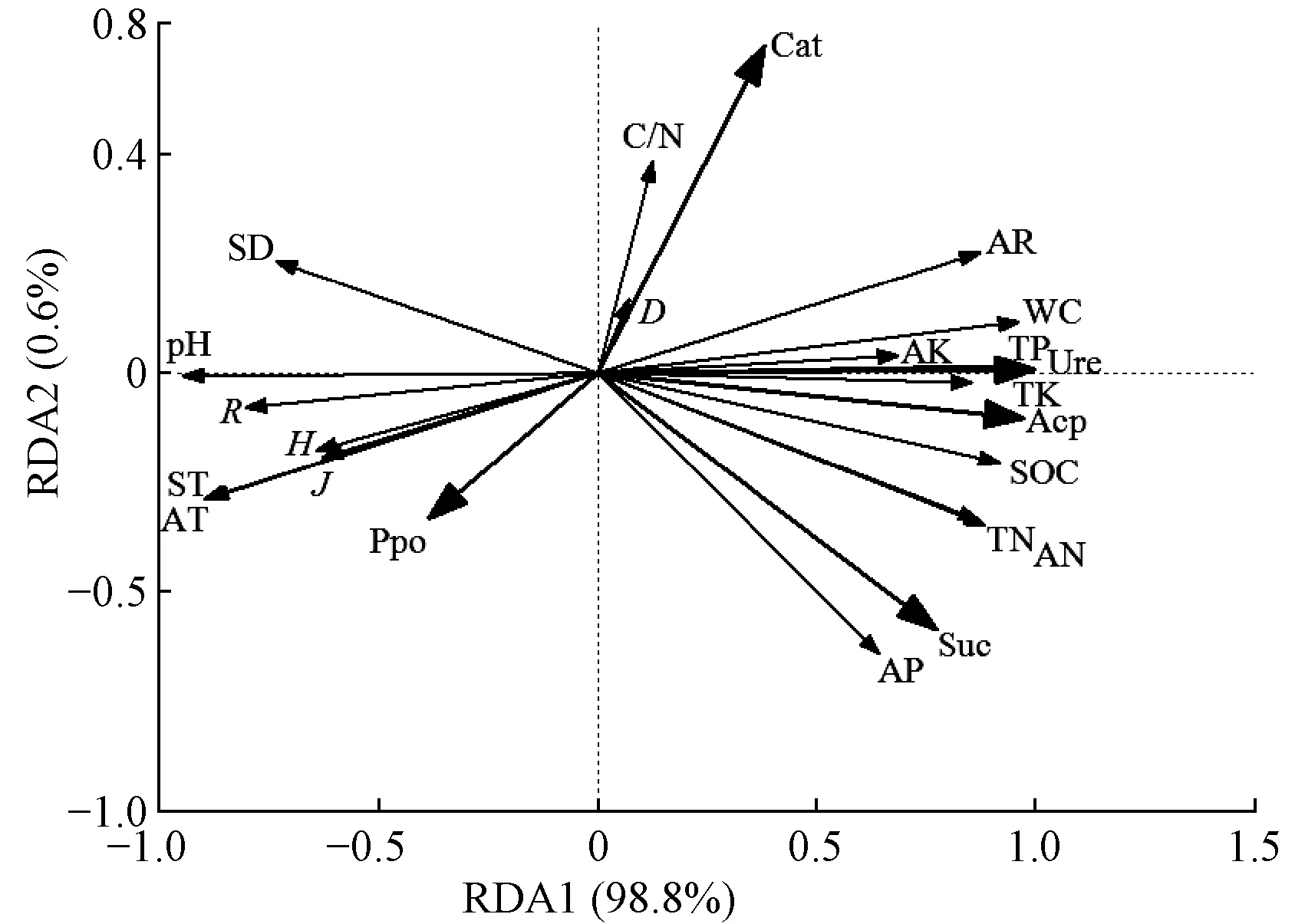
图2 土壤酶活性与环境因子的冗余度分析
综上可知,不同环境因子对土壤酶活性的影响差异较大。进一步使用蒙特卡洛检验对影响酶活性的环境因子重要性进行排序,可得全磷含量>含水量>pH>有机碳含量>年均气温>土壤温度>水解性氮含量>年降水量>全氮含量>全钾含量>Margalef 丰富度指数>土壤密度>速效钾含量>有效磷含量>Shannon-Wiener 多样性指数>Pielou 均匀度指数>C/N>Simpson 指数(表6)。重要性排序1~10的各因子解释量明显高于其他因子。相关分析显示,除C/N和Simpson 指数对土壤酶活性影响较小且未达显著水平外,其余环境因子对土壤酶活性的影响均达到极显著水平(P<0.01)。
3.4.2 单一环境因子对土壤酶活性的影响 由图3a可知,全磷含量与过氧化氢酶、脲酶和酸性磷酸酶活性呈正相关,与多酚氧化酶和蔗糖酶活性呈负相关。由图3b可知,酸性磷酸酶活性箭头完全落入实践圈内,脲酶和蔗糖酶活性箭头大部分落入实践圈内,过氧化氢酶活性箭头部分落入虚线圈内,多酚氧化酶活性箭头未落入线圈内,说明土壤含水量对酸性磷酸酶活性影响较大,对脲酶和蔗糖酶活性影响次之,土壤含水量增减对过氧化氢酶活性具有抑制作用,而对多酚氧化酶活性无影响。由图3c可知,土壤pH与多酚氧化酶活性呈正相关,与过氧化氢酶、脲酶和酸性磷酸酶活性呈负相关。由图3d可知,多酚氧化酶和蔗糖酶活性箭头落入有机碳的虚线圈内,而过氧化氢酶活性箭头完全落入实线圈内,脲酶和酸性磷酸酶活性箭头部分落入实线圈内,说明有机碳含量与过氧化氢酶活性呈极显著正相关,对多酚氧化酶和蔗糖酶活性起抑制作用。由图3e、g和j可知,年均气温、水解性氮含量和全钾含量与多酚氧化酶、蔗糖酶、脲酶和酸性磷酸酶4种酶活性呈正相关,与过氧化氢酶活性呈显著负相关。由图3f可知,土壤温度与多酚氧化酶活性呈正相关,与过氧化氢酶、脲酶、酸性磷酸酶和蔗糖酶活性呈负相关,表明土壤温度仅对多酚氧化酶活性具有促进作用。由图3h可知,年均降水量与多酚氧化酶和蔗糖酶活性呈正相关,与过氧化氢酶活性呈负相关。由图3i可知,全氮与多酚氧化酶和蔗糖酶活性呈正相关,与脲酶、酸性磷酸酶和过氧化氢酶活性呈负相关。由图3k可知,Margalef丰富度指数与脲酶、酸性磷酸酶和蔗糖酶活性呈负相关。由图3l可知,土壤密度与过氧化氢酶活性呈正相关,与多酚氧化酶和蔗糖酶活性呈负相关。

表5 土壤酶活性RDA排序的特征值及累积解释量

表6 环境因子变量解释的重要性排序和显著性检验结果
4 讨论
4.1 土壤酶活性对植物群络多样性及水热条件的响应
海拔梯度变化是决定生境差异的主导因子,继而制约着植物群落的空间分布格局(马克平等,1995),植物种类和组成不同,其根系分泌物和枯落物的质和量就不同,从而可能引起土壤酶活性差异(杨万勤等,2001)。本研究表明,3种海拔梯度植被带土壤的脲酶、蔗糖酶和酸性磷酸酶活性表现为中山湿性常绿阔叶林>半常绿季雨林>沟谷雨林,与谷晓楠等(2017)、曹瑞等(2016)研究结果类似。中山湿性常绿阔叶林海拔较高,地表枯枝落叶大量积累,土壤表层累积大量养分,虽然土壤温度较低,但降水量却很大,地表枯落物使土壤温度保持相对稳定,为土壤酶的分解作用提供了一个稳定环境,而群落指数与土壤酶活性呈不同程度负相关,可能与土壤脲酶、蔗糖酶和酸性磷酸酶对分解底物具有专一性有关;沟谷雨林虽然植被群落组成丰富,其枯枝落叶多,但养分分解和流失较快,土壤酶活性可能会在短时间内较高,但得不到长期维持,因此其含量反而相对减少。本研究也表明,土壤脲酶、蔗糖酶、酸性磷酸酶和过氧化氢酶活性与年均降水量呈极显著正相关,说明充足的土壤水分能为各种酶促反应提供较好条件,从而使酶活性随土壤含水量升高而增加(Jinetal.,2009)。
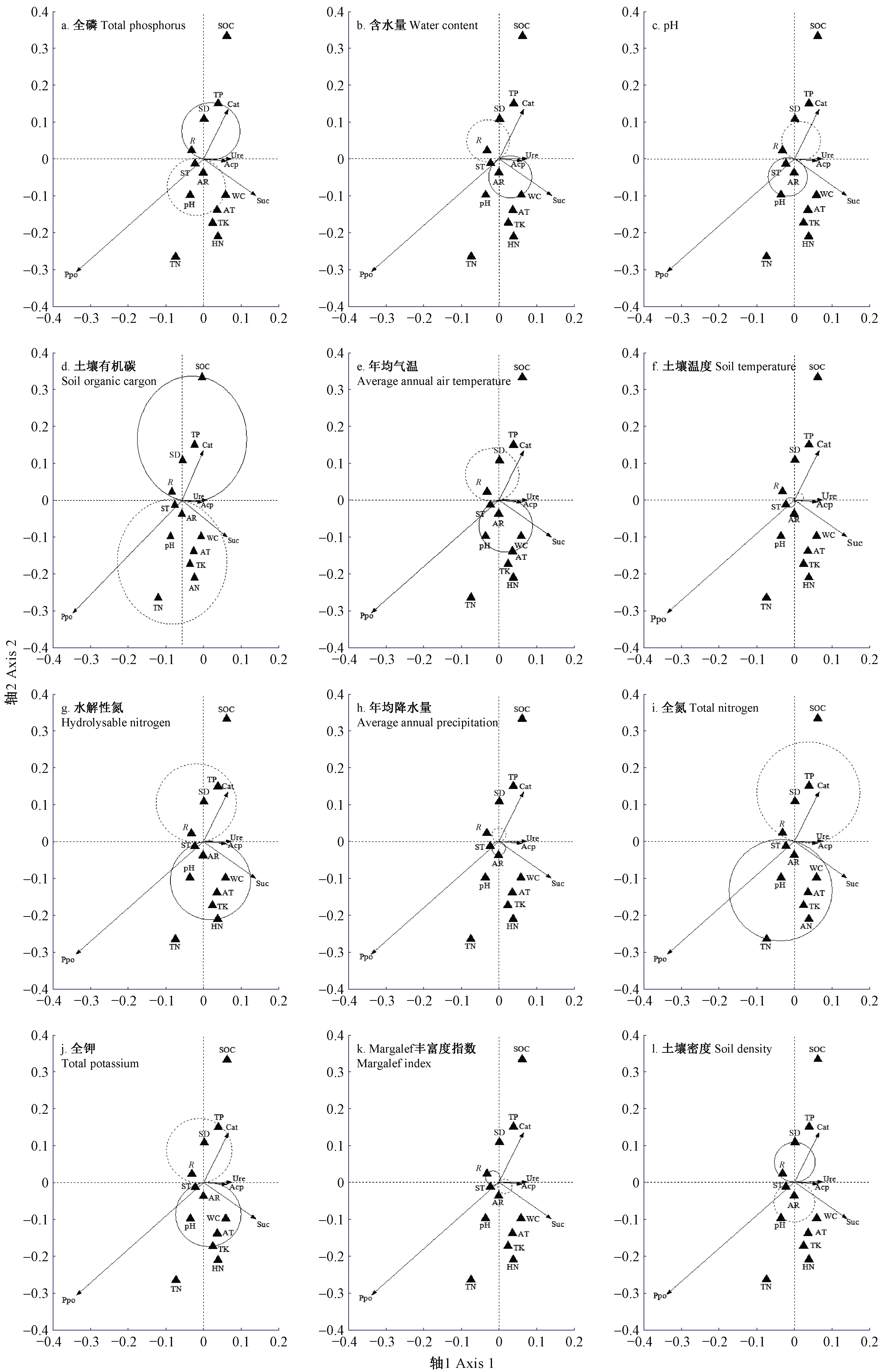
图3 单一环境因子对土壤酶活性影响的检验结果
通常认为,在一定温度范围内,酶活性与土壤温度呈正相关(Kochetal.,2007)。本研究表明,年均气温和土壤温度与脲酶、蔗糖酶、酸性磷酸酶和过氧化氢酶活性呈显著负相关,可能原因是温度升高会增加土壤蒸发量,降低土壤水分含量,进而限制土壤酶活性;而年均气温和土壤温度与多酚氧化酶活性呈正相关。不同海拔梯度植被带土壤温度和年均气温差异较大,说明温度是控制酶活性的重要因素之一,升高或降低温度对提高酶活性均有显著影响。本研究中土壤多酚氧化酶活性与物种多样性指数(Margalef 丰富度指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数)及水热条件表现出不同程度的正相关,说明沟谷雨林凋落物中可能含有大量酚类物质,使沟谷雨林土壤多酚氧化酶活性升高,而其他土壤酶活性相对降低。研究表明,当土壤中水溶性酚含量升高时,多酚氧化酶活性也随之增强(杨梅等,2012)。不同植物群落结构会形成独特的土壤微气候,进而改变土壤动物和微生物的营养源,这种关系又影响到土壤生物多样性(Hooperetal.,2000)、土壤物理结构、化学组成和生物活性(Griersonetal.,2000),使得不同区域内的土壤酶活性出现差异。因此,随海拔升高,温度降低,植被物种多样性降低,促进了特定种类土壤酶活性;降水量增加,可能抵消了温度下降产生的限制作用,进而使得土壤酶活性随海拔升高而增加。
4.2 土壤酶活性对土壤理化性质的响应
一般认为,土壤有机碳、全氮等养分含量增加可促进酶活性(荆瑞勇等,2015)。本研究表明:土壤脲酶、蔗糖酶和酸性磷酸酶活性与土壤全磷含量、含水量、有机碳含量、水解性氮含量、全氮含量和全钾含量呈显著正相关,与土壤密度和pH值呈极显著负相关,说明这3种酶活性与土壤碳、氮、磷素的积累和循环有关。谷晓楠等(2017)研究表明,在高海拔处大量有机质积累,养分含量高,为土壤微生物的新陈代谢提供了大量分解底物,使得土壤酶活性增加。此外,含水量增加直接影响土壤中的酶促作用和生物化学反应强度,从而影响土壤酶活性(解丽娜等,2014;Jinetal.,2009)。土壤密度直接影响土壤中养分、水分、空气和微生物等的迁移通道、贮存库和活动场地,还影响植物生长及微生物细胞的代谢过程,进而影响土壤酶活性。
本研究发现,单一环境因子对5种酶活性的影响存在一定差异,这可能与各种酶的特性不同有关。土壤脲酶活性体现了土壤中N转化强度,蔗糖酶活性高低取决于土壤中有机质含量和C/N的高低,土壤磷酸酶活性高低直接影响着土壤中有机磷的分解转化及其生物有效性,过氧化氢酶活性与土壤有机质转化速度密切相关(周礼凯,1987);而土壤多酚氧化酶与土壤有机质的形成有关(贾新民等,1995),土壤含氮量大小直接影响土壤中的水解酶和氧化酶活性(张艺等,2017)。本研究表明,土壤有机碳含量、全氮含量、水解性氮含量、含水量、全钾含量、全磷含量和pH值这7个单一环境因子对酶活性的提高具有重要作用。有机碳及氮能增加植被地上及地下根系生物量,促进根际微生物繁殖,可能促进多酚氧化酶、蔗糖酶、脲酶和酸性磷酸酶活性增加。含水量与脲酶、蔗糖酶和磷酸酶活性呈正相关,可能对此3种酶活性的提高具有促进作用。此外,土壤磷、钾素也能促进植物根系生长,增强土壤微生物及根系分泌土壤酶,说明土壤全钾、全磷含量增加利于提高过氧化氢酶、脲酶和酸性磷酸酶的活性,全钾含量增加还有利于提高多酚氧化酶和蔗糖酶的活性。土壤pH值过高或过低将直接影响植物的生长和微生物活动,从而影响土壤酶活性。本研究中,脲酶、蔗糖酶、酸性磷酸酶和过氧化氢酶与pH值显著负相关,表明pH值可能是抑制土壤酶活性的化学指标之一。这与陈双林等(2010)和何东进等(2017)的研究结果不同,可能由不同地区间土壤理化性质的异质性造成,因为土壤酶活性不仅受土壤pH值影响,还受到其他养分元素含量的影响。
本研究中,随海拔升高,各植被带土壤过氧化氢酶活性和多酚氧化酶活性与理化性质间的相关性不同于其他3种酶,可能与不同海拔梯度植被带的枯落物构成及根系分泌物不同有关。研究表明,凋落物中的木质素可以通过某种途径刺激土壤中过氧化氢类物质产生,使土壤生物加速分泌过氧化氢酶来抵抗氧化胁迫(郑学昊等,2018),从而使各植被带土壤过氧化氢酶出现差异。而土壤多酚氧化酶则是主要的木质素分解酶,其能氧化酚类化合物中的苯环,也对腐殖质的合成与分解具有重要作用(Sinsabaugh,2010)。杨承栋等(1995)研究发现,土壤多酚氧化酶活性与真菌多样性呈极显著正相关,可见,土壤有机质大量积累会激活过氧化氢酶和多酚氧化酶活性(张咏梅等,2004)。但也有研究表明,氮含量增加对土壤过氧化氢酶没有显著影响(Freyetal.,2004)。Li等(2014)发现,无机氮增加会抑制多酚氧化酶活性,而有机氮加入会降低这种抑制作用。因此,为能更进一步理解土壤多酚氧化酶和过氧化氢酶与土壤理化性质间的相关性,还需不断探究与土壤微生物间的机制关系。
4.3 土壤酶活性沿海拔梯度的变化
本研究表明,随海拔升高,各植被带土壤酶活性存在差异,其中土壤脲酶、蔗糖酶、酸性磷酸酶和过氧化氢酶活性均表现为中山湿性常绿阔叶林>半常绿季雨林>沟谷雨林。3种不同海拔梯度植被带土壤含水量、土壤温度和土壤养分含量差异各不相同,与土壤脲酶、蔗糖酶、酸性磷酸酶和过氧化氢酶活性在3种不同海拔梯度植被带间的变化趋势一致,说明海拔梯度变化会引起土壤水分和热量的重新分配,改变植被群落构成,使土壤养分含量在不同海拔梯度出现差异,导致土壤酶活性出现明显变化。不同海拔梯度植被带土壤脲酶、蔗糖酶、酸性磷酸酶和多酚氧化酶活性均随土层加深而降低,表现出显著的表聚特性,与Khknen等(2011)的结果相似。这可能是由于土壤表层累积了较多枯枝落物,腐殖质含量较高、养分含量高、细根生物量高、土壤疏松、水热条件和通气状况良好,有利于土壤微生物生长,土壤微生物代谢活跃,使表层的土壤酶活性增强(Bardelliabetal.,2017;Loeppmannetal.,2016)。随着土层加深,密度增大、根系分布减少、土壤水热条件恶化,限制了土壤微生物正常活动,进而影响土壤微生物的代谢产酶能力,这些因素的综合作用使得土壤酶活性随着土层加深逐渐降低(张仕艳等,2010;Jangetal.,2010)。综上所述,海拔梯度变化主要引起植被群落构成的变化,进一步引起土壤理化性质变化,进而调控土壤酶活性。
5 结论
随海拔升高,水热条件和植被群落多样性表现出不同变化趋势。年均降水量呈增加趋势,年均气温和土壤温度呈下降趋势;Simpson 指数呈增加趋势,Margalef 丰富度指数、Shannon-Wiener 多样性指数和Pielou 均匀度指数呈下降趋势。土壤温度、水分变化促进了植物群落结构变化,使得不同海拔梯度土壤酶活性出现显著差异。
随海拔升高,土壤理化性质表现出不同变化趋势。土壤含水量及有机碳、全氮、全磷、全钾、水解性氮、有效磷和速效钾含量呈上升趋势,土壤密度和pH呈降低趋势;土壤C/N呈先增后降趋势,且对不同土壤酶活性产生促进或抑制作用。冗余分析表明,土壤全磷、含水量、pH值和有机碳是调控土壤酶活性的主要因子;单一环境因子中的土壤磷、水分含量、pH值、碳、氮和钾含量对土壤酶活性的强弱起重要作用。
南滚河国家级自然保护区内各植被带土壤酶活性存在显著的海拔差异,其中土壤脲酶、蔗糖酶、酸性磷酸酶和过氧化氢酶活性均表现为中山湿性常绿阔叶林>半常绿季雨林>沟谷雨林,而土壤多酚氧化酶活性表现为沟谷雨林>半常绿季雨林>中山湿性常绿阔叶林;除过氧化氢酶活性在不同海拔梯度植被带均随土层加深而增加外,其余土壤酶活性均随土层加深而降低。
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26 00:44:12
生物技术通报(2020年12期)2020-12-21 09:20:00
腐植酸(2020年1期)2020-11-29 00:15:44
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07 11:14:32
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:45
广西林业科学(2016年1期)2016-03-20 05:33:00
中国粮油学报(2016年5期)2016-01-23 02:44:40
医学研究杂志(2015年8期)2015-06-22 14:00:57
茶叶通讯(2014年2期)2014-02-27 07:55:39
中国医学科学院学报(2013年6期)2013-03-11 20:26:04
