
贝壳沙埋深度对杠柳幼苗生长及光合特性的影响
2020-06-02 10:13孙景宽刘京涛杨红军宋爱云
江西农业大学学报 2020年2期
李 田,孙景宽,刘京涛,杨红军,宋爱云
(滨州学院 山东省黄河三角洲生态环境重点实验室,山东 滨州256603)
【研究意义】黄河三角洲贝壳堤属于典型的古海岸地貌,为粉沙淤泥质海岸带,其基质成分主要包括海生贝壳及其碎片、细沙、淤泥质粘土等[1]。沉积物在贝壳堤的形成和发展过程中具有极其重要的作用[1]。泥沙的沉积速率不仅影响贝壳堤的形成速度,还会影响其盐度以及植被受潮汐淹没的程度;而泥沙掩埋被认为是影响种子萌发、出苗、幼苗存活、苗木生长以及盐分变化的重要因素,从而对植物栖息地的稳定性产生严重影响[2]。因此在海岸带退化植被恢复重建中,研究沙埋胁迫对植物的影响及其适应机制具有重要的实践指导意义。【前人研究进展】由于风、潮汐以及人类活动的强烈影响,贝壳堤上的植物经常受到贝壳沙掩埋的威胁。泥沙掩埋是影响贝壳堤生态系统植被分布和组成的重要因素。植物通过泥沙运动和沉积作用进行掩埋,并进化出各种适应方式,从而允许其在相对紧张的可变条件下进行生存、生长和繁殖[3-4]。但不同的掩埋深度对植物生长等方面会产生不同的影响,从而改变着植物的稳定性和脆弱性[5-6]。相关研究已证实许多沙丘植物可以较好的适应这种泥沙掩埋胁迫,有些植物则需要在适当的沙埋条件下才能维持更好的生长状况[7-9]。这是由于不同掩埋深度会改变土壤基质的一些理化性状,其中适当埋深可降低土壤温度和增加湿度,利于种子的萌发以及植物成长;但掩埋过深时,又会造成光照减少、土壤透气性降低等不利条件。此外,植物对泥沙掩埋沉积物的响应不同还取决于它们的形态和生物量的分配差异[10]。以往关于沙埋对植物影响的研究主要包括沙埋对种子萌发及其存活率的影响,沙埋对植物形态特征、生物量分配的影响,以及沙埋对植物抗逆生理响应等方面[11-15],但目前有关沙埋对植物光合特性影响的研究报道不多[16-17],且上述研究主要集中在干旱或半干旱的沙漠或沙丘地区,而有关海岸带贝壳堤的相关研究甚少。【本研究切入点】杠柳(Periploca sepium Bunge)是黄河三角洲贝壳堤海岸带典型的优势灌木,具有防风固沙等多种抗逆特性。目前仅见有关于干旱或盐胁迫下对杠柳种子萌发、生理特性及光合作用特征的研究[18-22],而沙埋作为贝壳堤区域一种常见且严重的环境胁迫因素,影响和限制着该区域的植物生长与发育,但目前有关贝壳堤沙埋对杠柳幼苗生长及光合特性的研究尚未见报道,对贝壳堤区域沙埋因素对植物的影响研究也显然不足。因此本研究拟对不同深度贝壳沙埋深下杠柳幼苗的生长和光合特性进行研究,探讨杠柳幼苗对贝壳沙埋深度的耐受极限,明确贝壳沙埋深与杠柳幼苗生物量及其光合特性参数间的相互作用关系。【拟解决的关键问题】本研究旨在探讨杠柳幼苗生长及光合特性对贝壳沙埋深度的响应规律,解析杠柳在植物生物量和光合特性方面适应贝壳沙掩埋的相关生长和生理策略,为贝壳堤海岸带植被的恢复重建提供科学依据。
1 材料与方法
1.1 试验材料
试验所用的杠柳种子及贝壳沙于2016年10月取自滨州贝壳堤岛与湿地国家级自然保护区,其中贝壳沙含盐量0.05%。采集后的种子晾晒风干后,密封4℃保存。
1.2 试验设计
将杠柳种子均匀点播在装有贝壳沙的塑料盆中。种子在播种时埋深为0.5 cm,每盆播种30粒,每个处理3次重复,共计15盆,于数控温室中自然光照培养。培养30d后选取植株大小基本一致的幼苗进行定苗,每盆定苗10株,之后用贝壳沙小心进行掩埋,贝壳沙埋深度分别为幼苗高度的0%、33.3%、66.7%、100%、120%。
1.3 测定项目及方法
埋深处理后每隔15d统计植株的成活率;60d后控制叶室内温度25℃,CO2390µmol/mol,应用CID-340型光合作用系统测定顶端生长点下方的第3片叶片的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci),每个处理测定3张叶片。另外计算下列指标:瞬时水分利用效率(W U E)=Pn/Tr;表观光能利用效率(LUE)=Pn/P AR;羧化效率(CE)=Pn/Ci。同时测量株高和根长及根、茎、叶的干质量。
1.4 数据分析
采用SPSS17.0进行单因素方差分析和Duncan多重比较。选用结构方程模型(structural equation modeling,SEM)分析埋深、生物量和植物光合特性参数间的相互作用关系,其中结构方程模型构建采用AMOS 17.0.2软件进行,数据检验采用maximum likelihood(ML)法,渐进残差均方和平方根(root mean square error of approximation,RMSEA)指标用于评价模型的适配度。
2 结果与分析
2.1 不同贝壳沙埋深对杠柳幼苗存活率的影响
贝壳沙埋深33.3%和66.7%的处理及对照在整个试验阶段均保持了极高的成活率(图1)。埋深15d时,各处理及对照间的成活率差异均不显著,但随着处理时间的延长,埋深100%和120%的处理成活率均有所降低,且显著低于其他处理和对照。埋深120%的处理较100%处理的成活率下降趋势更为明显,特别是120%处理下杠柳植株在埋深60d后几乎全部死亡,而100%埋深处理的杠柳幼苗成活率仍可达到53.3%。
2.2 不同贝壳沙埋深对杠柳幼苗生长指标的影响

图1 不同贝壳沙埋深下杠柳植株的存活率Tab.1 Survival rate of Periploca s epium Bunge buried in different depth of shell sand
不同贝壳沙埋深对杠柳幼苗株高和根长的影响如表1所示。由于埋深达到120%时,杠柳幼苗无法存活,因此仅对0%、33.3%、66.7%、100%沙埋处理下的杠柳幼苗相关指标进行了分析。从表1可以看出,各处理杠柳幼苗株高和根长均随着贝壳沙埋深的增加先增加后降低。其中,埋深33.3%的杠柳幼苗株高和根长生长最好,分别高于对照55.2%和51.1%,其次是埋深66.7%的杠柳幼苗,分别高于对照32%、15.9%,埋深100%的株高和根长分别低于对照49.6%、43.4%。统计分析表明,杠柳株高各处理和对照及各处理间差异均显著,杠柳根长除了埋深66.7%和对照差异不显著外,其他处理和对照及各处理间差异均显著(P<0.05)。这一结果表明,轻度埋深(33.3%)能够促进杠柳幼苗的长高及其根系的生长,而完全埋深(100%)对其株高和根系生长均起到显著的抑制作用。
不同贝壳沙埋深对杠柳幼苗根、茎、叶干质量具有显著性影响(P<0.05)。其结果如表1所示,与对照相比,杠柳根、茎、叶干质量均随着贝壳沙埋深的增加呈现先增加后下降的变化趋势。其中埋深33.3%时,杠柳叶、根、茎各部分干质量较对照及其他处理均表现出明显优势,分别高于对照51.4%、50.0%、91.7%;其次是埋深66.7%时,杠柳各部分干质量也均高于对照,但其中叶干质量和根干质量与对照差异不显著;对于埋深100%时,根茎叶各部分干质量值最低,分别低于对照50.0%、25.0%和43.2%,其中除茎干质量与对照差异不显著外,其余部分干质量均显著低于对照及其他处理。进一步对比在不同沙埋深度下,杠柳幼苗根、茎、叶干质量的变化幅度存在差异,其中轻度埋深下(33.3%),变化幅度由大到小表现为:茎、叶和根;中度埋深下(66.7%),由大到小依次为,茎、根和叶;完全埋深下(100%)由大到小依次为,根、叶和茎。
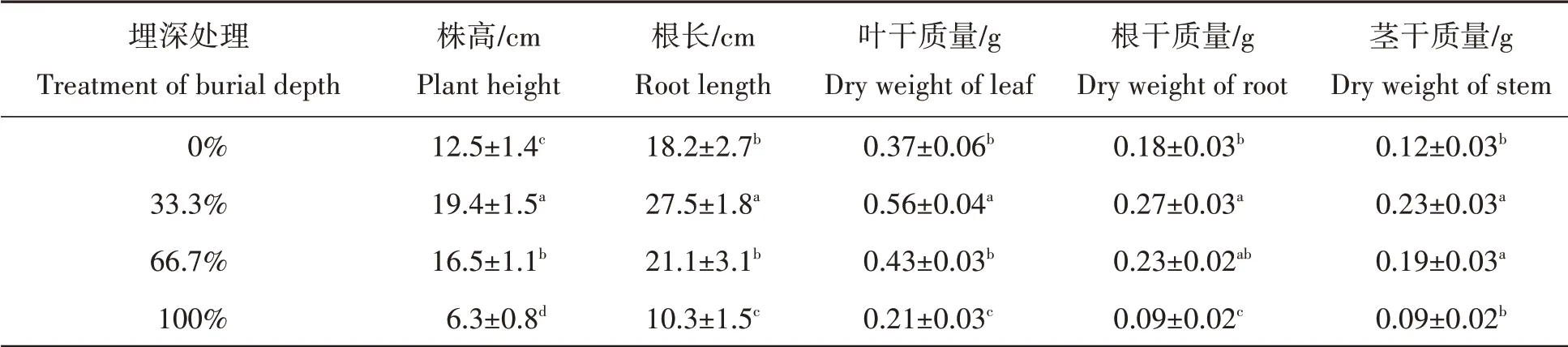
表1 不同贝壳沙埋深对杠柳幼苗生长指标的影响Tab.1 Effects of different sediment depth on growth indices of Peri pl oca se pium Bunge seedlings
2.3 不同贝壳沙埋深对杠柳幼苗生物量结构的影响
从图2可以看出,贝壳沙埋深下杠柳的生物量结构发生了明显变化。其中沙埋深33.3%时,杠柳根的比重较对照略微下降,沙埋深66.7%时较对照增加,沙埋深100%时有所降低,但各处理和对照间的根占比未达到差异显著水平(P>0.05)。随着沙埋深度的增加,杠柳叶比重除完全埋深外,均有所降低,但各处理及对照间差异不显著;茎比重则相反呈增加趋势,其中埋深66.7%时茎占比最大,与其他各处理间差异不显著,但显著高于对照(P<0.05)。

图2 不同贝壳沙埋深对杠柳生物量构成的影响Fig.2 Effects of different sediment depth on the biomass composition of Peri ploca s epium Bunge seedlings
2.4 不同贝壳沙埋深对杠柳幼苗光合特性的影响
不同贝壳沙埋深对杠柳幼苗光合特性的影响如图3所示。其中,各处理下杠柳净光合速率随着贝壳沙埋深的增加呈先增加后降低趋势。沙埋深33.3%、66.7%时,杠柳幼苗净光合速率分别高于对照73.1%、54.0%,但两者之间差异不显著;沙埋深100%时高于对照31.0%,但与对照未达到差异显著水平。整体来看,随埋深深度的增加,杠柳的净光合速率值仍维持在较高水平,在所设定的埋深范围内即使完全埋深条件下,杠柳的Pn值仍略高于对照,并未出现显著的光抑制现象,表明杠柳可承受较深深度的沙埋处理。各处理下杠柳蒸腾速率Tr随着贝壳沙埋深的增加均有不同程度的增加,其中埋深0%时,杠柳幼苗蒸腾速率最低;埋深66.7%、100%时,Tr值均显著增大,且分别显著高于对照13.7%和25.5%(P<0.05);33.3%埋深下杠柳幼苗蒸腾速率达到最大,显著高于对照40.5%(P<0.05)。上述结果表明,即使是完全埋深下,杠柳叶片的蒸腾作用并未受到明显抑制,且在一定范围内,随着埋深深度的增加,蒸腾速率反而呈现增加趋势,提示埋深深度的增加并未引起杠柳水分的亏缺,其原因可能在于较深的埋深增强了土壤中的相对含水量。
不同沙埋处理下,杠柳幼苗的胞间CO2浓度随着贝壳沙埋深的增加呈微弱的上升趋势,其中,埋深33.3%和66.7%时,杠柳叶片胞间CO2浓度较对照分别提高1.9%和5.4%;在埋深深度100%时杠柳Ci值达到最大,比对照提高5.9%;但各处理及其对照之间未达到差异显著水平(P>0.05)。进一步对不同贝壳沙埋深下杠柳幼苗的羧化效率C E进行分析,结果显示,各处理下杠柳羧化效率均随着贝壳沙埋深的增加而有不同程度的增加;埋深33.3%、66.7%、100%时,羧化效率分别高于对照71.2%、45.5%、24.2%(P<0.05),其中33.3%和66.7%埋深下二者差异不显著,完全埋深处理和对照间差异不显著。说明不同埋深环境下杠柳叶肉细胞对CO2的亲合能力存在差异,从而影响杠柳叶片进行羧化反应的效率。

图3 不同贝壳沙埋深对杠柳光合特性的影响Fig.3 Effects of different sediment depth on the photosynthetic characteristics of Pe ri pl oca sepium Bunge seedlings
不同贝壳沙埋深对杠柳幼苗光能利用效率和水分利用效率的影响显示,不同贝壳沙埋深下杠柳幼苗光能利用效率随着贝壳沙埋深的增加呈先增加后降低的趋势,但沙埋深100%时仍高于对照。其中随着埋深深度的依次增加,杠柳光能利用效率分别显著高于对照66.7%、54.1%、31.5%(P<0.05)。统计分析表明,33.3%和66.7%埋深时,杠柳光能利用效率差异不显著,66.7%和100%沙埋深时,差异不显著;其余处理间及对照间均差异显著。环境条件的差异会引起叶片光能利用效率的变化,本研究中光能利用效率的响应趋势与净光合速率的变化情况相一致,即在不同埋深深度下,杠柳仍保持着较高的光能利用效率,表明其对沙埋的适应能力较强。不同贝壳沙埋深对杠柳幼苗水分利用效率的影响显示,各处理下杠柳水分利用效率的变化趋势与光能利用效率不同。随着埋深深度的增加,杠柳水分利用效率先升高后降低,在沙埋深66.7%时,杠柳水分利用效率最高,显著高于对照34.0%,但与沙埋深33.3%时差异不显著;完全埋深时,杠柳水分利用效率有所降低,但仍与对照无显著差异。结果表明,沙埋深33.3%时就可达到杠柳水分利用效率的较高值,继续增加埋深深度对提高杠柳水分利用效率作用不显著。
2.5 埋深处理下杠柳生长指标与光合特性的相关性分析
从表2可以看出,不同沙埋深度下,杠柳的株高和根长与各部分的生物量显著相关,且达到极显著水平(P<0.01),在光合测定指标中,净光合速率(Pn)、水分利用效率(WUE)、光能利用效率(LUE)和羧化效率(CE)均与株高、根长以及茎干质量间存在显著相关性(P<0.05)。不同沙埋程度下杠柳资源利用效率与Pn、Tr之间存在密切相关性,其中水分利用效率(W U E)与Pn呈显著正相关,且达到极显著水平(P<0.01);光能利用效率(L U E)与Pn、Tr和W UE均呈显著正相关,羧化效率(C E)与Pn、Tr、WUE和L U E均呈显著正相关(P<0.01)。
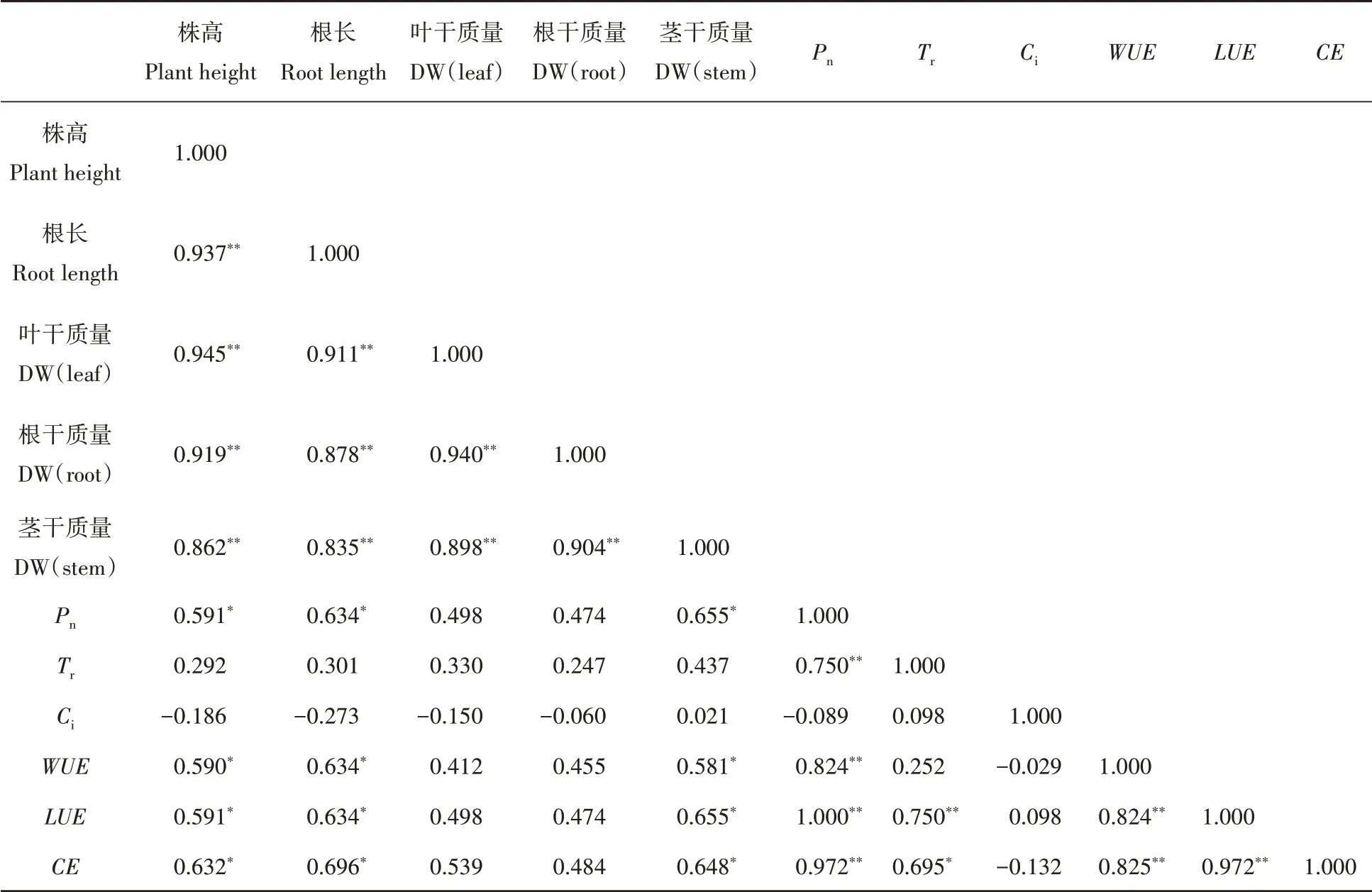
表2 杠柳生长指标与光合特性间的相关性分析Tab.2 Correlation between growth index and photosynthetic characteristics of Peri ploca sepium Bunge
2.6 埋深处理下杠柳生长及光合特性的结构方程模型分析
为更好的明确不同埋深处理与杠柳生长及光合特性参数间的关系,笔者对相关性分析结果中具有显著性关联的指标进一步采用了结构方程模型对它们之间的作用效应进行了分析,对它们之间的直接和间接效应予以解释(图4)。结构方程模型分析显示,模型中RMSEA值均小于0.05,表明模型适配度良好。
分析结果表明,沙埋处理对杠柳株高和根长的直接效应均达到差异极显著水平(P<0.001),且均为负效应,效应系数分别为-0.57和-0.90(图4A,C),但沙埋对茎干质量的直接效应差异并不显著(P>0.05)(图4B);从各项光合参数来看,其中净光合速率对株高和茎干质量的影响最大,为正效应,两者效应系数分别为0.87和0.91,且均达到极显著水平(P<0.001)(图4A,B)。蒸腾速率对根长的直接效应不显著(P>0.05),而水分利用效率对根长的直接作用效应达到极显著水平,效应系数达0.61(P<0.001)(图4C)。结构方程模型分析显示,净光合速率和水分利用效率可直接对杠柳株高、根长、茎干质量产生正效应,且相应的效应系数显示两者对后者具有较大的作用效应,这进一步说明了,在一定埋深范围内,杠柳株高、根长、茎干质量的增加主要是由杠柳净光合速率和水分利用效率的增强引起的,这与杠柳生长指标与光合特性因子间的相关性分析结果相一致。
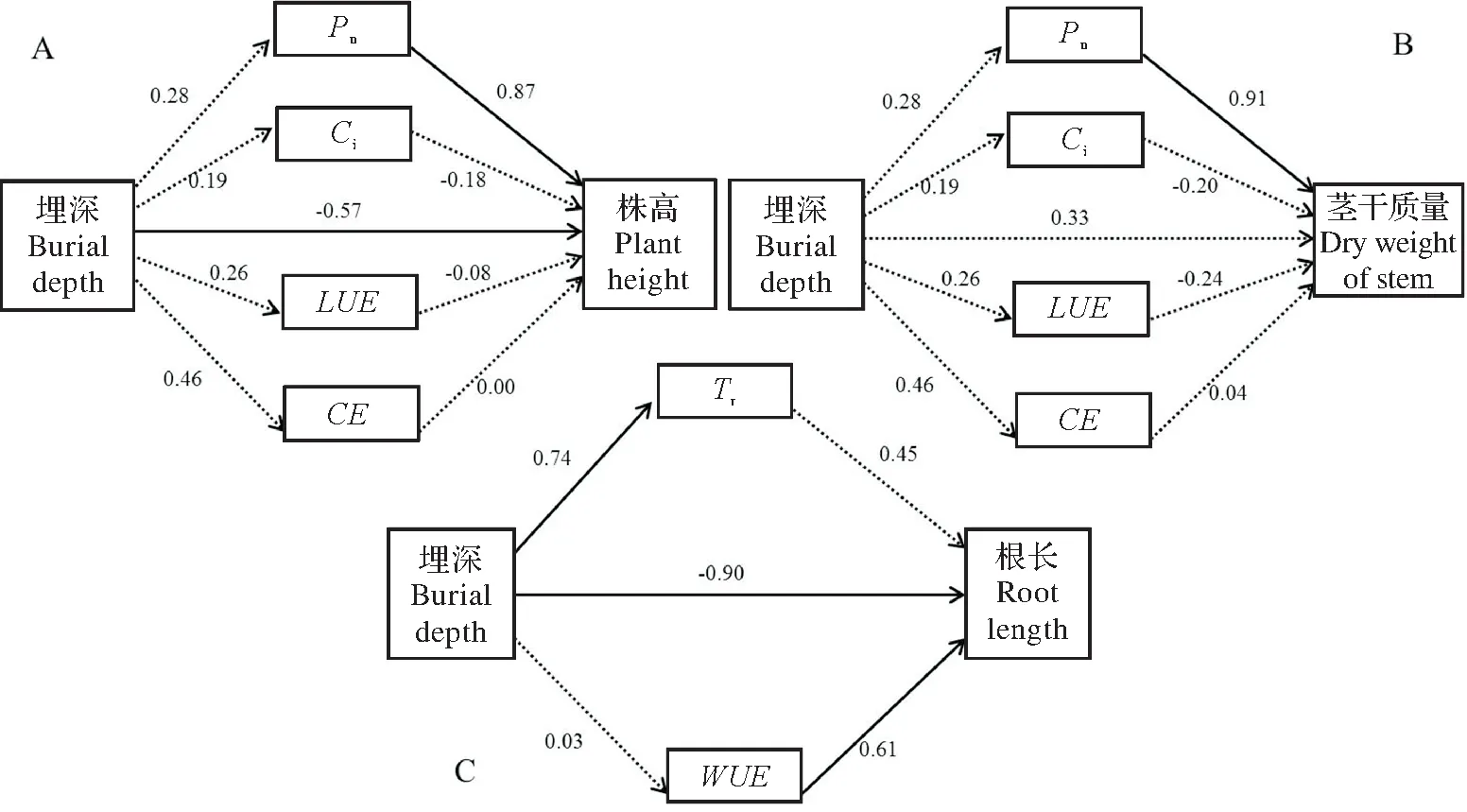
图4 贝壳沙埋深与杠柳生长及光合参数的结构方程模型分析Fig.4 Analysis of shell sand burial depth,growth and photosynthetic parameters of Peri pl oca sepium Bunge with structural equations
3 结论与讨论
不同植物对沙埋深度的耐受极限不同,研究植物的埋深阈值响应对于筛选固沙植物具有重要的实践指导意义。通常,不同的埋深度对植物的存活和生长具有不同的影响,但当埋深超过其耐受阈值时,则会抑制植物的生长甚至造成死亡[23-24]。本研究发现杠柳在完全埋深时仍能存活,所能忍受的最深埋深阈值深度可达100%;另一方面,杠柳对不同埋深度的阈值反应具有时间性,不同埋深度下的杠柳存活率随着埋深时间的延长,埋深度越高(100%和120%),其死亡率越高,且两者与适当埋深(33.3%和66.7%)下的杠柳成活率差异也越明显。从杠柳埋深的耐受深度阈值来看,其明显强于无芒雀麦、砂生槐、小叶锦鸡儿、梭梭[25-28],与羊柴、盐地碱蓬以及分布于海滨沙地的砂引草这类可耐全埋的植物相近[13,24,29],但同时又低于耐沙埋能力更高的沙生植物沙米[30]。这表明,植物的耐掩埋能力存在很大差异,而杠柳在完全掩埋后,在长达60d的埋深后仍能破土生长,保持一定的存活率,说明其耐沙埋能力较强,这也是杠柳能够成为贝壳堤岛上优势物种的主要原因之一。但要保证杠柳的正常存活与生长,其对沙埋深度仍存在严格的界限要求,即埋深度应不超过杠柳的埋深阈值(<株高的120%)。其原因在于过度沙埋造成沙土阻力过大,幼芽出土困难而不能保证光合作用,且长时间的呼吸消耗致使植株能量耗尽而死亡。这也是目前杠柳仅适用于在固定或半固定沙地地区固沙绿化,而不能生长在流动沙丘上的主要限制因素,因此在这些流动沙丘地区,可采用一定的人工保护措施来提高杠柳的存活率。
对于生长在潮汐地附近的植物,能够忍受一定程度的沙土掩埋是这些地区优势植物物种得以生存的必要条件。杠柳作为贝壳堤岛上的主要优势物种,在生长和生理方面形成了适应沙土掩埋的相关策略。其中资源的分配和利用是植物最基本和最重要的生命活动,不同植物种类,其资源分配策略不同[31]。当植物被掩埋时,一些沙丘植物个体会将生物量和养分等资源从根部转移到地上部分,以此保持其光合能力[32],而某些沙丘植物在相同的情况下其根冠比却没有发生明显变化[33],有些则相反,其地下部分的增加大于地上部分[34]。从沙埋下杠柳生长特性来看,杠柳对沙埋的形态响应在某种程度上类似于遮阴环境下植物对光的竞争[35-36]。结合不同贝壳沙埋深下杠柳生物量结构来看,茎比重的增加最为明显,这表明在应对沙埋胁迫中,杠柳植株将更多的资源用于茎的生长,且随埋深度的增加,其茎占比也不同程度的增大。这是由于沙埋可导致杠柳幼苗对光源产生竞争,引起杠柳在垂直结构上的生长变化,如杠柳茎的伸长,导致茎干质量和株高的增加,从而保证了杠柳幼苗从沙土里顺利钻出,以便获得更多的光照。
沙埋通常被认为是限制沙地或者沙漠植物存活与生长发育的重要因子[37]。以往很多关于埋深下植物的生长适应性研究主要是基于对生物量的测定,而对埋深下植物的生理反应,特别是光合作用反应的研究很少[25,38]。本研究发现,沙埋对杠柳幼苗的光合特性影响显著,其所表现出的光合作用特点与沙埋下沙米、骆驼刺、樟子松Pn、Tr等光合参数增加的结论具有一定的相似性[30,39-40]。可见,在适应沙埋环境中,通过增强光合特性来保证自身的正常生长,是许多耐沙埋植物普遍采取的生理策略[41]。沙埋之所以会影响植物的光合特性,其原因在于植物生命活动中存在着重要而基本的资源分配与利用[42]。研究[43]表明,适当的埋深可促使植物提高地上组织叶片中的叶绿素成分和N含量,这一适应性调整对幼苗植株增强其光合作用能力具有重要作用。不同深度贝壳沙埋深下杠柳幼苗光合能力的增强,是杠柳适应沙埋胁迫的一种积极表现,这是由于沙埋减少了植物总的光合面积,从而启动了受影响植物的补偿响应过程,即较高的净光合速率,从而弥补减少的光合面积,平衡植物对碳等物质和能量资源的需求[44-45]。此外,杠柳在完全埋深较长时间后,其光合特性与已报道的某些植物在完全埋深时的表现存在很大差异,如研究[16]发现差巴噶蒿的Pn、Tr、Gs随沙埋深度的增加显著降低,且完全埋深下其下降趋势更明显。造成上述差异的原因可能在于,不同植物类型其耐沙埋能力不同,杠柳在完全埋深期间,顶部不断有新的嫩叶长出,进而使光合作用得到有效保障,表明其耐沙埋能力较强。再次,不同沙埋深度的增加可导致土壤湿度相对增加,因此在一定程度上了缓解和避免了光抑制现象的发生,从而使杠柳光能利用率提高[46]。
结构方程模型(SEM)是一种强大的、多变量技术,用于检验和评估多变量间的因果关系,能够明确给出各关系的大小;该模型可以对预先假定的因果关系的直接和间接影响进行检测,是将因子分析和路径分析相结合的高级统计方法[47]。SE M作为强大的分析工具,为生物学领域的相关研究提供了极其有利的技术手段,如利用该模型分析油菜各个性状之间的因果关系大小,用以评价土壤化学与原核生物群落组成之间的关系等[48-49],这些已建立的SE M模型均较好的说明了相关问题。针对本研究中要揭示的埋深处理、杠柳生长以及光合特性三者间所存在的具体作用效应问题,笔者构建了符合适配度要求的结构方程模型,得到了较相关性分析更为深入的结果:特别是在所分析的各项光合参数中,笔者发现沙埋处理仅对杠柳蒸腾速率产生了极显著的直接效应。依据相关研究,Tr可影响植物的Pn、Gs和WU E,其中Tr与WU E存在显著的正相关性,因此较强的蒸腾速率有利于增加植物的水分利用效率[50]。因此,笔者可以得到一条明确的路径关系,即埋深处理通过直接影响杠柳的蒸腾速率进而间接影响其水分利用效率,而水分利用效率又直接影响了植物生长指标中的根长。此外,从效应大小来看,虽然埋深对根长产生了直接显著的负效应,但其效应系数远远小于水分利用效率对杠柳根长的直接正效应,这在一定程度上揭示了沙埋能够促进植物生长的原因,这是由于相关的研究表明,沙埋深度的增加可以促进土壤含水量的增加,有利于植物水分利用效率的提高[30]。这与植物受到干旱、高温等胁迫后的水分利用效率机理不同,上述胁迫往往能引起植物水分亏缺,造成植物Pn和Tr的降低,从而迫使植物在缺水状态下通过增强水分利用效率保证光合作用的有效进行[51]。而在本研究中,沙埋下杠柳的蒸腾速率并没有降低反而增加,这是由于蒸腾速率一般随土壤水分的加大而增强,说明沙埋对于杠柳保持有效的水分利用效率是有利的。
猜你喜欢
今日农业(2021年10期)2021-11-27
趣味(语文)(2021年12期)2021-04-19
小哥白尼(野生动物)(2019年6期)2019-10-10
今日农业(2019年14期)2019-01-04
读友·少年文学(清雅版)(2018年4期)2018-01-14
学苑创造·A版(2016年5期)2016-06-21
中国火炬(2015年12期)2015-07-31
植物营养与肥料学报(2014年1期)2014-03-11
中国火炬(2013年3期)2013-07-24
植物营养与肥料学报(2012年1期)2012-10-26
