
烟草祖先种及重要野生种PDR1基因的生物信息学分析
2020-05-18 08:57:16平文丽李雪君孙计平孙焕
中国烟草学报 2020年2期
平文丽,李雪君,孙计平,孙焕
河南省农业科学院烟草研究所/烟草行业黄淮烟区烟草病虫害绿色防控重点实验室,河南许昌 461000
植物的多向耐药性(Pleiotropic drug resistance,PDR)转运蛋白是ATP 结合盒(ATP-binding cassette,ABC)转运蛋白家族的G亚家族蛋白[1]。ATP转运蛋白广泛存在于原核、真核生物的细胞膜系统中,参与多种初生代谢物、次生代谢物的跨膜运输,但PDR所属的亚家族只存在于植物与真菌中。植物的PDR蛋白广泛参与到营养吸收、发育调控、解毒过程以及植物对生物胁迫和非生物胁迫的防御反应中[2,3]。近年来,针对模式植物拟南芥和主要作物的研究还表明,PDR是多抗基因,与多种植物对真菌病原的抗性相关,例如,AtPDR12参与拟南芥对真菌病害的抗性[4-5];水稻中的OsPDR9[6]及大豆中的GmPDR12均具有抗真菌的作用[7]。从小麦中分离到与镰刀菌抗性相关的两个PDR类基因TaPDR1和TaPDR7,能通过Ca2+信号转导相关途径及跨膜转运由镰刀菌产生的代谢毒素脱氧雪腐镰刀菌烯醇(Deoxynivalenol,简称DON毒素)等方式,减轻病菌对小麦的毒害[8-10]。在烟草研究中,目前已经证实渐窄叶烟草(Nicotiana attenuata)中的NaPDR1和NaPDR1-like基因对真菌类病原链格孢菌(Alternaria alternata)的抗性至关重要[11];普通烟草中的NtPDR1基因参与烟草对病原的抗性反应中[12];蓝茉莉叶烟草(Nicotiana plumbaginifolia)中ABC家族的NpABC1基因参与次生代谢物萜类物质的分泌,所以与真菌抗病性相关[13],但关于普通烟草PDR基因与真菌病害抗性的关系尚鲜见报道。
烟草镰刀菌根腐病是由镰刀菌引起的一种真菌性病害,会导致烟草品质下降、产量降低、烟草生产产值降低。近年来该病害在我国河南、湖南、福建等各大烟区均有发生,且发生情况呈上升趋势[14]。防治该病害危害最经济有效的手段是选育抗病品种,但是目前普通烟草中关于抗镰刀菌根腐病的种质资源、相关抗病基因的报道较少、已知具有抗镰刀菌根腐病的烟草种质资源较少,抗病育种工作有赖于挖掘栽培烟草祖先种、野生近缘种中所蕴含的抗病基因。普通烟草(Nicotiana tabacum)基因组(SSTT)中S基组及T基组分别来源于祖先种林烟草(S)、绒毛状烟草(T)。野生资源是指除了两个栽培种(普通烟草、黄花烟)之外的所有烟草属的植物种[15]。这些植物种是在长期的自然选择条件下保留下来的类型,其中某些重要的野生种中蕴含着丰富的抗病基因,是烟草育种工作中抗病基因主要来源。如黏烟草(N.glutinosa)是白粉病和普通花叶病的重要抗源,长花烟草(N.longiflora)和蓝茉莉叶烟草(N.plumbaginifolia)是黑胫病的重要抗源[15-16];但是对烟草镰刀菌根腐病的抗源、抗病基因的研究仍鲜见报道。前人的研究表明,PDR基因具有广谱抗病性,且与小麦等植物对镰刀菌属病原的抗性相关,基于此,本文从相关数据库中提取烟草及其祖先种和重要近缘野生种的PDR基因和蛋白序列,采用生物信息学方法对其结构、功能等特征进行分析和预测,以期为后续的基因功能研究、基因编辑、抗病分子育种等工作提供科学依据和理论基础。
1 材料与方法
1.1 材料
选择的研究对象包括:普通烟草(Nicotiana tabacum)、烟草的祖先种林烟草(Nicotiana sylvestris)和绒毛状烟草(Nicotiana tomentosiformis)、重要野生烟草本氏烟(Nicotiana benthamiana)、蓝茉莉叶烟草(Nicotiana plumbaginifolia)、渐窄叶烟草(Nicotiana attenuata)等的DNA和蛋白序列。
1.2 方法
依据模式植物拟南芥中与抗真菌相关的AtPDR12基因的蛋白、普通烟草NtPDR1基因序列,通过BLAST比对等方式,从拟南芥数据库TAIR(https://www.arabidopsis.org/)[17]、NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)、本氏烟数据库Benthgenome(http://benthgenome.qut.edu.au/)[18-19]、 茄 科 植 物 数据 库 solgenomics(https://solgenomics.net/organism/Nicotiana_attenuata/genome)[20]等多个数据库中查询烟草祖先种及重要近缘野生种的PDR1基因;通过ExPASy网站的翻译工具(Translate,https://web.expasy.org/translate/)翻译为PDR1蛋白序列、采用蛋白参数分析工具(ProtParam,https://web.expasy.org/protparam/,以及 protscale:https://web.expasy.org/protscale/)分析这些基因的理化特性;用TMHMMM(http://www.cbs.dtu.dk/services/TMHMM/)、pepwheel(http://emboss.bioinformatics.nl/cgi-bin/emboss/pepwheel)、phyre2(www.sbg.bio.ic.ac.uk/phyre) 等工具预测其跨膜结构、蛋白结构;用SUBA4(http://suba.live/)[21]在线分析相关蛋白在细胞中的定位;用比较基因组学工具CoGe Blast(https://genomevolution.org/coge/CoGeBlast.pl#)分析普通烟草各条染色体上PDR1基因的高度相似的序列;用BDGP(https://fruitfly.org/seq_tools/promoter.html)分析基因的转录起始位点、用PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) 和 PLACE(https://www.dna.affrc.go.jp/PLACE/)等工具预测NbPDR1基因启动子区的顺式作用元件,进而进行功能预测;最后,用 CRISPRdirect(https://crispr.dbcls.jp)[22]在 线 工 具预测了NtPDR1基因的基因编辑位点。
2 结果与分析
2.1 普通烟草、烟草祖先种、重要近缘野生种中PDR1蛋白的理化特征
以AtPDR12序列为探针,通过序列比对(TBLASTN和BLASTP等)从各个数据库中获取烟草、祖先种及近缘野生种的PDR基因如下:普通烟草中有5个相似序列:NtPDR1,NtPDR2,NtPDR3,NtPDR4及NtPDR5;林烟草、绒毛状烟草、本氏烟、渐窄叶烟草中均有3个相似序列:PDR1、PDR2和PDR3;蓝茉莉叶烟草中有3个相似序列PDR1,PDR2和PDR5。在其他已测序的野生烟草,如黏烟草(Nicotiana glutinosa,绒毛烟草组,2n=24)、福尔吉特氏烟草(Nicotiana forgetiana,花烟草组,2n=18)、花烟草(Nicotiana alata,花烟草组,2n=18)、浅波烟草(Nicotiana repanda,浅波烟草组,2n=48)的基因组中未搜到相似序列。综合文献资料中烟草属的分类信息,黏烟草和普通烟草分别属于普通烟亚属的不同组别;福尔吉特氏烟草、花烟草、浅波烟草均属于碧冬烟亚属的花烟草或浅波烟草组,和普通烟草分别属于不同的亚属。推测这些烟草属植物虽然具有相同的祖先,但在进化过程中染色体数目、结构发生了变异,导致这些物种中某些基因结构与普通烟草有较大差别或发生了基因丢失。
对所得序列按照得分(Score)高、比对覆盖度(query coverage)高、与AtPDR12序列一致程度(Percent identity)高(高于60%)的标准进行筛选,得到的均为PDR1基因,所以本文选择各物种中的PDR1基因、蛋白作为研究对象。
利用ExPASY网站上的工具软件对普通烟草、祖先种及其重要近缘野生种的PDR蛋白的氨基酸序列及其理化特性进行了预测分析。结果表明,几个不同烟草的PDR1蛋白的分子量均与拟南芥相类似,为161-162 kD;组成该蛋白的氨基酸数为1432-1436个,其中组成跨膜区的氨基酸数为289-293个,略多于拟南芥;烟草中PDR1蛋白的等电点为6.3-6.56,比拟南芥ATPDR12的等电点低;蛋白的半衰期为30小时,不稳定指数介于33.07至35.81之间,是在植物中稳定存在的蛋白。各蛋白的分子式及其具体特征参数如表1所示。

表1 烟草及其祖先种、主要近缘野生种中PDR1蛋白的特征参数Tab.1 Characteristic parameters of PDR1 in tobacco and its ancestral species and major wild species
2.2 PDR1蛋白氨基酸序列的比对及结构域分析
针对模式植物拟南芥中AtPDR12基因及其编码的蛋白研究表明,PDR编码ABC转运蛋白(ATP-结合盒转运蛋白),该基因在拟南芥的叶、花、气孔、根等组织中均有表达,编码的蛋白包含跨膜结构域及核苷酸结合结构域,其亚细胞定位为细胞质膜上(SUBA data)[21]。序列比对结果表明,烟草祖先种、多种近缘野生种均存在ABC转运蛋白的保守功能域NBD和TMD,功能域之间靠疏水结构连接,各个模序(motif)序列(包括Walker A、Q-loop、ABC标 签、Pro-loop、Walker B、D-loop、H-loop等)与拟南芥等物种中PDR序列及排列顺序完全一致[23],暗示烟草中PDR蛋白完全符合PDR蛋白的特征、该蛋白的结构域与其功能密切相关,在进化过程中非常保守、无任何突变。具体结构域及序列如图1所示。
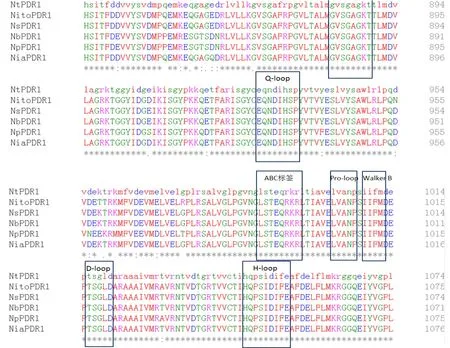
图1 烟草及其祖先种、主要近缘野生种中PDR1蛋白序列比对及在进化中保守的蛋白结构域(其中方框为ATP结合区域里的几个motif)Fig.1 Sequence alignment of PDR1 proteins in tobacco and its ancestral species, major wild species and conserved protein domains in evolution, where the boxes indicate motifs in the ATP binding area
根据在线工具TMHMM的分析,本文研究的普通烟草、烟草祖先种及重要近缘野生种中所有的PDR1蛋白均由2个核酸结合区(nucleotide-binding domains,NBD)和2个分别由6-7个跨膜α螺旋(membrane spanning alpha helices)构成的跨膜区(Transmembrane domain, TMD)组成。从N端到C端,依此为NBD1,TMD1,NBD2和TMD2。其中NBD1位于蛋白的N端,NBD2位于蛋白的中部,跨膜区TMD1和TMD2位于蛋白的中部和C端,见图2。该特征与拟南芥等其他物种中PDR蛋白的结构域特征也完全相符[23]。
本研究运用在线工具pepwheel对几种烟草PDR1蛋白的NBD和TMD之间的氨基酸序列进行了分析,得出的轮状图见图3。结果表明:在该区域中由疏水氨基酸和亲水氨基酸组成了螺旋状的疏水结构,符合ABC转运蛋白的结构特征。
2.3 NtPDR1蛋白的3级结构预测
综合Expasy网站提供的在线工具phyre2等蛋白三级结构工具的预测结果,以普通烟草的NtPDR1为例,本文分析了普通烟草NtPDR1蛋白的三级结构。结果表明,该蛋白包含了14个嵌入细胞膜的区段,N端和C端则在膜外侧。左侧的7个螺旋状结构域组成核酸结构域NBD;右侧的α螺旋组成跨膜区TMD。两者之间靠氨基酸的疏水作用连接,三级结构模型如图4所示。
2.4 比较基因组学分析
采用CoGe Blast分析普通烟草基因组各条染色体上PDR1基因的相似序列。结果如图5所示。在烟草的第9、13、14、15及23条染色体上,均有与PDR1基因高度相似的序列,CoGe BLAST结果显示,这五条染色体上相似序列的E值分别为4e-170,3e-117,0,0,2e-100,根据生物信息学原则,E值小于10-6即视为序列高度相似,因而推测这些染色体上均有与PDR1同源的序列;其中第14、15条染色体上的序列的E值等于0,相似度最高,推测PDR1基因即位于第14、15号染色体上。其他几个染色体上的序列有高达70%的部分与NtPDR1完全一致。该结果暗示PDR基因在生长发育或应对胁迫中有重要作用,因而在基因组里有冗余。

图2 烟草及祖先种、重要野生种中PDR1蛋白的跨膜结构域分析Fig.2 Transmembrane domain analysis of PDR1 protein in tobacco, its ancestral and important wild species
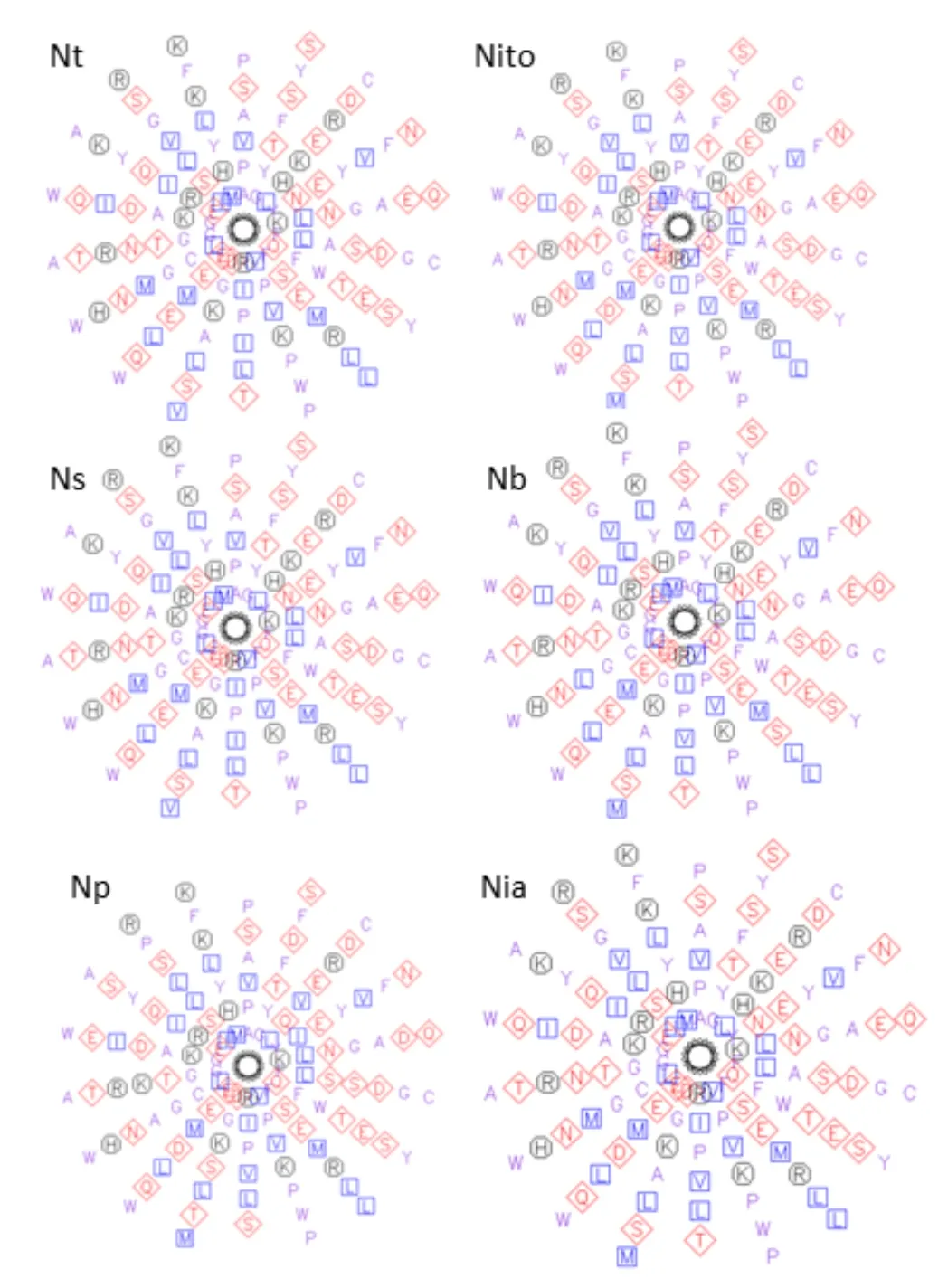
图3 烟草祖先种及重要野生种的PDR1蛋白中NBD与TMD之间的疏水氨基酸结构预测Fig.3 Prediction of hydrophobic amino acid structure between NBD and TMD domain of PDR1 in tobacco, its ancestors and important wild species
2.5 启动子的顺式作用元件预测
植物基因组中与抗病、抗逆相关基因一般仅在特定的情况下表达,以避免给植物正常生长带来的负面效应[24]。其表达调控主要是通过转录因子与基因启动子区域的顺式作用元件的结合,启动、增强或者抑制基因的表达,从而调控下游基因表达、抵御外界胁迫。本文从数据库中提取NbPDR1基因上游的3000bp的DNA序列,利用BDGP分析该基因的转录起始位点,结果表明,只有1个转录起始位点,启动子所在位置为554-604 bp处。其启动子序列为:CA AACTATTATATATAGAGAGAGGTGACAAAGAAAA AAGGAAAAGAGTGT。

图4 NtPDR1蛋白的3级结构。图中颜色为彩虹色,从红到蓝紫色指示蛋白从N端到C端的氨基酸结构Fig.4 The tertiary structure model of NBD and TMD of NtPDR1 in Nicotiana tabacum.The amino acid structure of the indicator protein from the N-terminal to the C-terminal ranges from red to blue-purple
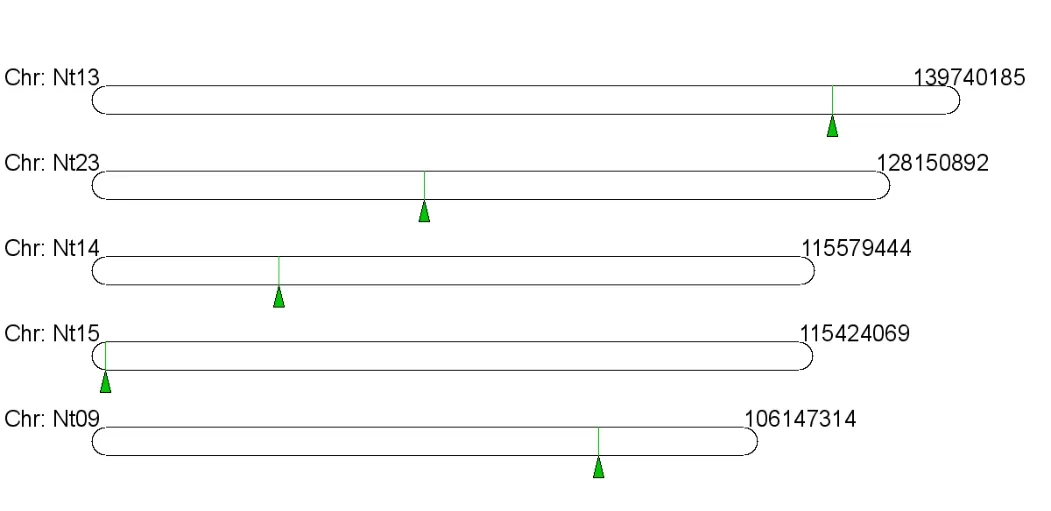
图5 普通烟草基因组上与NtPDR1高度相似片段的定位Fig.5 Locations of highly similar fragments of NtPDR1 on the tobacco (Nicotiana tabacum) genome
用Plant Care和PLACE预测了NbPDR1的启动子区的顺式作用元件。综合对比对其结果,得出的顺式作用元件如表2所示。对顺式作用元件进行分析,得出以下结论:NbPDR1基因启动子的调控元件主要包括4类:1)组成型元件,如31个TATA-box、39个CAAT-box等;2)病原菌诱导、与抗病相关的响应元件,其中包括1个真菌诱导响应元件ELRECOREPCRP1、3个抗真菌病害相关的元件BIHD1OS、以及其他多个病原真菌响应相关的元件;其中,启动子区还有一个DON毒素诱导表达的特异元件-300ELEMENT,表明该基因受到镰刀菌的诱导后表达上调,暗示该基因可能参与DON毒素的细胞转运、解毒相关、能够减轻镰刀菌及其毒素对烟草的毒害,与烟草耐DON毒素、对镰刀菌引起的病害的抗性相关。3)大量与植物抗病防御中起到关键作用的激素的应答等相关的元件,其中包括35个与水杨酸(SA)诱导相关的元件GT1CONSENSUS、32个与脱落酸(ABA)干旱胁迫响应相关的元件MYCCONSENSUSAT、1个与茉莉酸(JA)响应相关的元件CGTCA-motif;4)其他,包括1个钙离子响应元件ABRERATCAL。

表2 NbPDR1启动子的顺式作用元件预测结果Tab.2 Prediction of cis-acting elements in promoter regions of NbPDR1 in Nicotiana benthamiana
2.6 基因编辑位点分析
采用在线gRNA设计网站CRISPR direct,以NGG为PAM序列,设计针对普通烟草NtPDR1的基因编辑靶位点。得到的CRISPR/Cas9基因编辑的靶位点中,得分最高、脱靶概率最低的4个基因编辑位点信息如表3所示。这四个靶位点中,两个位于负义链(-),两个位于正义链(+),序列的GC含量适中,20 bp全长序列和PAM及12 bp种子序列和PAM的靶位点在整个基因组范围内都只有1个,脱靶概率较低,表明,该基因存在至少4个适用于基因编辑的靶位点。

表3 CRISPR/Cas9基因编辑靶位点Tab.3 Target site of CRISPR/Cas9 gene editing
3 讨论和结论
PDR蛋白是植物ABC转运蛋白G亚家族的成员,在多种植物中的研究结果表明,PDR蛋白参与植物对多种生物胁迫及非生物胁迫的响应过程[1-7]。目前在普通烟草中已经报道的NtPDR1参与次生代谢物如二萜、倍半萜烯萜等物质的转运,以及叶面的环二萜类等物质的分泌,调节烟草对生物胁迫及非生物胁迫的抗性[12]。渐窄叶烟草NaPDR1和NaPDR1-like基因与真菌类病害抗性相关[11];近年来,在烟草BY2悬浮细胞中的实验证实,NtPDR1既参与烟草的组成型抗性,也参与烟草的诱导抗性[3]。
本文利用拟南芥和普通烟草中PDR基因、蛋白序列为基础,获取并分析了普通烟草、烟草祖先种及近缘野生种的PDR1基因、蛋白序列及结构,分析结果表明这些序列之间均存在高度相似性、蛋白的结构均含有核苷酸结合结构域NBD和跨膜结构域TMD,且包含了所有保守基序,符合经典的PDR结构特征。
此外,NtPDR1基因的重要同源基因NbPDR1基因启动子区的调控元件包括DON毒素诱导的特异启动子,暗示该基因的表达受到镰刀菌的正向诱导、可能与烟草对镰刀菌的抗性相关;也包含大量病原菌诱导、病原真菌诱导响应等大量抗病响应相关的元件,表明该基因的表达受到病原菌、真菌胁迫、多种逆境胁迫的诱导;该基因上游还有大量与ABA、SA、JA等多种激素响应相关的元件,这些激素均参与调控植物抗性,暗示PDR基因在植物抗病反应中的表达水平会受到激素诱导的影响。另外,从调控元件预测结果推断,NbPDR1基因还参与植物的耐旱、耐盐、机械损伤、钙离子响应等多种生理过程,暗示烟草的PDR1基因除了对真菌病害具有抗性外,还能够响应多种胁迫因素或者受胁迫相关的激素的诱导表达、防御逆境,表明该基因可能参与烟草对包括镰刀菌病原在内的多种生物、非生物胁迫的响应,是一个广谱抗性相关基因,在分子育种、抗病育种等工作中有较好的应用潜力。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
作物学报(2022年2期)2022-11-06 12:11:02
中国烟草学报(2021年3期)2021-08-04 03:50:42
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
生命科学研究(2018年1期)2018-05-29 01:12:47
江苏农业科学(2017年15期)2018-02-06 07:17:42
上海农业学报(2017年3期)2017-04-10 12:39:14
上海农业学报(2017年3期)2017-04-10 12:39:10
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
天津科技大学学报(2016年1期)2016-02-28 16:59:45
