
红枣经济林不同植被覆盖土壤水分入渗特征
2020-05-13 09:07:54刘焕焕王改玲殷海善贺润平
灌溉排水学报 2020年4期
刘焕焕,王改玲*,殷海善,贺润平,
(1.山西农业大学 资源环境学院,山西 太谷 030801;2.山西省农业科学院 植物保护研究所,太原 030031;3.山西省农业科学院 农业资源与经济研究所,太原 030006)
0 引 言
入渗过程是水分自土表垂直进入土壤的过程,是降水、地表水、土壤水和地下水相互转化过程中的重要环节,决定着降水进入土壤的数量。土壤水分入渗性能是评价土壤水土保持能力和抗侵蚀能力的重要指标[1-2],直接影响降水再分配过程中地表径流与土壤储水量的大小。红枣产业是山西省重要的经济林产业,红枣栽培面积33.33×104hm2,年总产量达60×104t以上,是山区农民的主要经济来源之一。其中,吕梁山区是山西省最大的红枣生产区,红枣种植面积为13.33×104hm2,占全省红枣种植面积的40%。然而,吕梁山位于黄土高原北部,地形破碎、植被覆盖稀少,降雨少而集中,水分流失严重。如何促进水分入渗,提高降水资源的利用率,提高枣园生产力,是实现该区生态及经济协同发展的关键。
土壤入渗能力受土壤干湿程度、孔隙状况和地表覆盖等因素的影响。国内外研究表明,植被覆盖通过根系形成的大孔隙,以及枯枝落叶层形成的有机质改良土壤结构,进而提高土壤渗透能力[3-5];此外,植被还可以拦截降雨,减少径流,增加入渗[6-7]。目前,国内外学者已就不同植被类型、植被覆盖度土壤入渗性能进行大量研究[8-9,19]。果园生草栽培是20世纪40年代兴起的现代化果园土壤管理模式,目前已在美国、日本、法国等国家广泛推广。我国于1988年将果园生草制作为绿色果品生产主要技术措施在全国推广,生草栽培对果园土壤水分及果树生长、产量的影响也有较多报道。但截至目前,生草的果园面积不足10%。林下生草覆盖对土壤入渗的影响还未见报道。吕梁山枣园采用的管理方式主要有2种:第一,清耕,即除去地表所有覆盖物,包括杂草等;第二,林下种植大豆等粮食作物。通过双环法对比研究红枣经济林不同管理方式下土壤水分入渗特征,并运用模型对入渗过程进行模拟,以期为控制红枣经济林水土流失提供一定理论基础。
1 研究区概况
试验地位于山西省吕梁市临县庞家庄,地理位置为 110.532013°E,38.037807°N,海拔928±1 m,属于大陆性季风气候,多年平均气温9.4 ℃,多年均降雨466.7 mm,雨热同期,降雨主要集中在夏季。土壤为黄土母质上发育的黄绵土。按照典型性和代表性的原则,选择该地多年清耕作业的红枣经济林地进行试验,枣树树龄为6 a。设置有A清耕作业区;B自然生草区;C大豆种植区,3个处理,每个处理3次重复,共9各小区,小区大小为5 m×4 m。试验区地形坡度为 20o,半阳坡,试验采用随机区组设计。清耕作业区1 a内3次耕作,除去地表所有覆盖物,包括杂草;自然生草区不采取任何管理措施,任杂草自然生长;大豆种植区,大豆播种时间为每年的5月,播种方式为穴播,播种密度为12穴/m2,播种后定期拔除杂草。试验开始于2017年3月,2018年为试验的第2年,各个小区的布设与2017年保持一致,小区布设示意图见图1。入渗试验进行时对样地植被进行调查,样地基本情况见表1。

图1 不同植被覆盖区布设示意图Fig.1 Schematic diagram in different vegetation coverage areas

表1 不同植被覆盖区样地基本情况Table 1 Basic conditions in different vegetation coverage areas
2 材料与方法
2.1 野外双环入渗试验
采用双环法测定土壤水分入渗率,内环直径为20 cm,外环直径为35 cm,高30 cm,用橡皮锤将内环和外环均垂直打入土壤,深度为12 cm。开始测定时向内环和外环同时注水,并使水层深度始终保持8 cm,利用医用注射器随时往内环内注水,以此保持内环水位一定,测定单位时间内内环所加的水量。测定开始时,每5 min记录1次消耗的水量,直到入渗达到稳定,总入渗时间为120 min,进而计算土壤水分入渗速率,入渗率的计算式[10]为:

式中:f为入渗率(mm/min);v为单位时间加水量(mm3/min);a为内环面积(mm2),本试验中,a为31 415.926 mm2。选取土壤初始入渗速率、平均入渗速率和稳定入渗速率作为评价土壤入渗性能的基本指标,其中土壤初始入渗速率(mm/min)为最初入渗时土壤的入渗速率,本试验选择前5 min的平均入渗速率作为初始入渗速率;平均入渗速率(mm/min)为测定时段内各入渗速率的平均值;稳定入渗速率(mm/min)为入渗达到稳定时土壤水分的入渗速率。
2.2 土壤体积质量和孔隙度的测定
在3个不同植被覆盖区内用环刀采集原状土,同时采集混合土样,采样深度均为0~20 cm,采集后带回实验室内进行土壤体积质量、土壤总孔隙度、毛管孔隙度、非毛管孔隙度及土壤含水率的测定。其中,体积质量采用环刀法的测定,总孔隙度根据体积质量计算,计算公式为:土壤总孔隙度=[(土壤比重-土壤体积质量)/土壤比重]×100%;毛管孔隙度、非毛管孔隙度采用圆筒浸透法[11]的测定;含水率采用烘干法[12]的测定。入渗试验和体积质量、孔隙度及含水率的测定于2018年10月9日进行。
2.3 土壤入渗模型选择
本研究选择Philip模型、Horton模型、Kostiakov模型[13-14]对不同处理小区果园土壤入渗过程进行模型拟合,如表2所示。

表2 土壤水分入渗模型公式及参数说明Table 2 Soil water infiltration model formula and parameter description
2.4 数据处理
采用SPSS 21.0进行显著性检验,模型拟合采用Origin8.0,利用Excel 2010进行制图。
3 结果与分析
3.1 不同植被覆盖区土壤的物理特征
如表3所示,不同植被覆盖区0~20 cm土层土壤体积质量和孔隙度不同,具体表现为自然生草区紧实,体积质量最大,总孔隙度最小;清耕区相对疏松,体积质量最小,总孔隙度最大;大豆区则介于自然生草区和清耕区之间。从大小孔隙的分布情况看,自然生草区毛管孔隙度最小,非毛管孔隙度最大;大豆区则表现为毛管孔隙度最大,非毛管孔隙度最小,清耕区2种孔隙孔隙度介于自然生草区和大豆区之间。
清耕区 0~20 cm土层土壤初始含水率最高,为10.44%,自然生草区最低,为9.43%,大豆区则介于清耕区和自然生草区之间。不同植被覆盖区0~20 cm土层土壤饱和含水率为36.34%~39.22%。
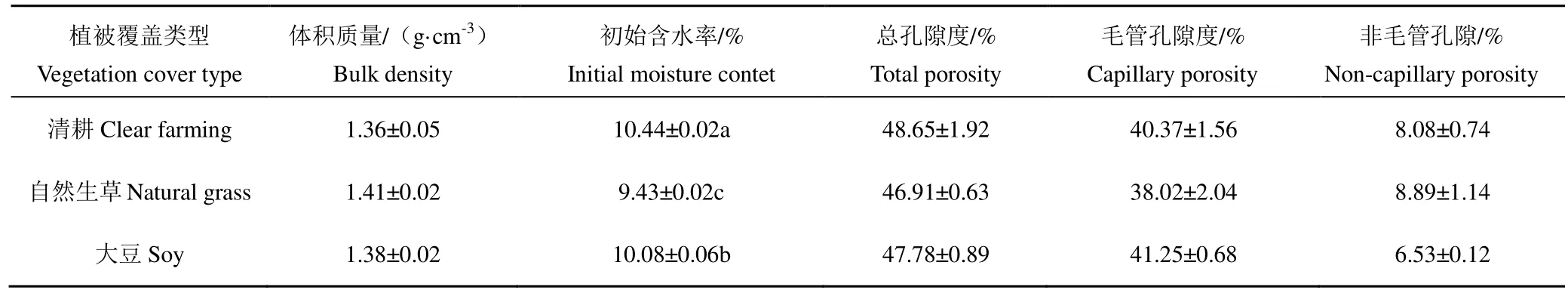
表3 不同植被覆盖区0~20 cm土层土壤物理性质Table 3 0~20 cm soil layer soil physical propertiesin different vegetation cover areas
3.2 不同植被覆盖区土壤水分入渗特征
不同植被区土壤入渗过程总体上可以分为 3个阶段(图2)。①入渗瞬变阶段(0~5 min):入渗初期土壤含水率较低,入渗水分在分子力的作用下,使下渗锋面快速延伸。此阶段内土壤入渗速率较高,入渗速率随时间急剧下降。②入渗渐变阶段(5~35 min),此阶段主要是水分在毛细管引力和重力作用下填充土壤孔隙过程,土壤入渗速率迅速降低。不同小区中,自然生草区入渗速率的减少速度明显快于其他 2种类型小区,至35 min时自然生草区与其他2种小区的入渗速率已基本相当。③稳定入渗阶段(35 min后),土壤孔隙被水分充满达到饱和后,水分在重力作用下向下渗透,入渗速率相对稳定,即为稳渗速率。
由表4可知,不同植被覆盖区土壤水分入渗速率具有相同的变化趋势,初始入渗速率最大,平均入渗速率次之,稳定入渗速率最小。3种小区土壤初始入渗速率表现为:自然生草区最大,为4.902 0 mm/min清耕区次之,为4.058 5 mm/min,大豆区最小,为4.055 3 mm/min。自然生草区土壤初始入渗速率明显高于清耕区、大豆区(p<0.05),而清耕区、大豆区差异不显著。稳定入渗速率为:清耕区最大,自然生草区次之,大豆区最小,其值依次为:1.952 8、1.743 5、1.559 7 mm/min,3种小区之间土壤稳定入渗速率均有显著差异(p<0.05)。平均入渗速率表现为:清耕区最大,大豆区次之,自然生草区最小,且清耕区与自然生草区、大豆区土壤的平均入渗速率差异明显,而自然生草区与大豆区差异不明显。

图2 不同植被覆盖区土壤水分入渗过程Fig.2 Soil water infiltration process in different vegetation cover areas
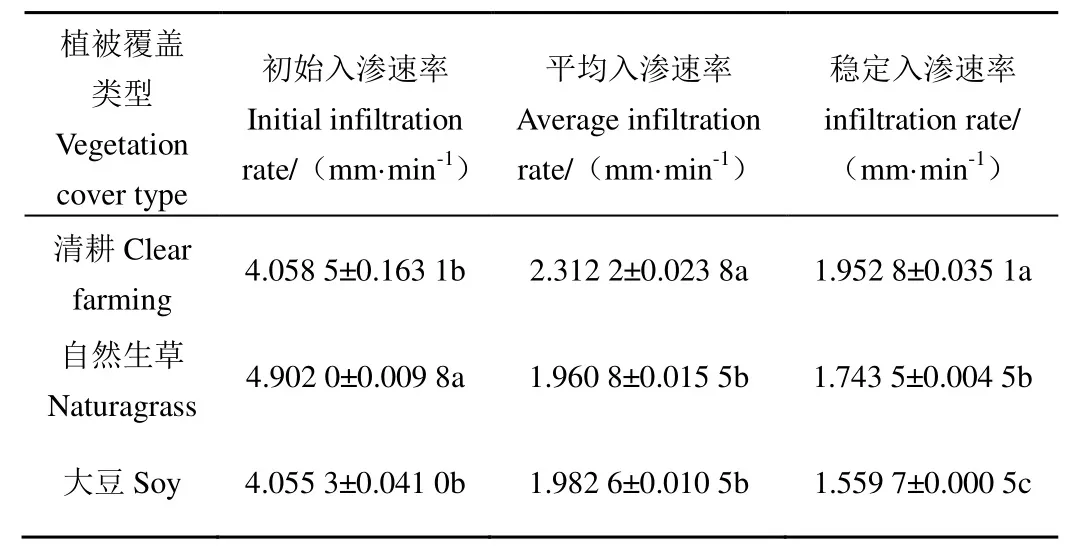
表4 不同植被覆盖区土壤入渗特征Table 4 Characteristics of soil infiltration in different vegetation cover areas
3.3 土壤入渗模拟分析
采用Philip模型、Horton模型和Kostiakov模型对不同植被覆盖区土壤水分入渗过程进行拟合,比较不同模型的拟合优度(表5)。
Philip模型中,S表示土壤吸渗率,反映了土壤入渗能力的大小,S值越大,表示土壤入渗能力越强。从拟合结果来看,S值大小依次为大豆区最大,其次为自然生草区,清耕区最小,与实测值存在一定偏差。土壤稳定入渗速率fc清耕区最大,其次是自然生草区,大豆区最小,与实测值大小顺序一致,但与实测值存在一定偏差。
Horton模型中,f0代表初始入渗速率,清耕区与大豆区土壤初始入渗速率分别为 4.543 mm/min,4.594 mm/min与实测值相差极小。土壤稳定入渗速率fc清耕区最大,为2.006 mm/min,其次是自然生草区,为1.796 mm/min,大豆区最小,为1.592 mm/min,其拟合结果与实测值大小顺序一致且偏差较小。
Kostiakov 模型中,参数a为经验入渗系数,a的主要影响因素有土壤含水率、结构、质地 和有机质等[16],a值的大小与初始入渗速率呈正比。清耕区、自然生草区、大豆区的a值分别为5.667、6.102、6.624与实测初始入渗速率的大小不一致,说明拟合结果与实测存在一定偏差。
Philip模型、Horton模型和Kostiakov 模型的决定系数R2分别介于为:从决定系数上来看和各参数所反映的结果来看,Horton模型对土壤入渗过程拟合效果最好,Philip模型次之,Kostiakov效果较差。
4 讨 论
土壤水分入渗是水分循环的一个重要环节,决定了降水转化为土壤水以及径流水的大小,直接影响降水资源的利用率。初始入渗速率、稳定入渗速率以及平均入渗速率是反映土壤入渗能力的常用指标。其中,表层土壤的初始入渗率与超渗产流密切相关。本试验结果表明,自然生草区土壤初始入渗速率明显高于清耕区和大豆区,这与自然生草区植被覆盖度大(60%)、土壤的初始含水率低有关。受草本植物根系影响,土壤大孔隙(非毛管孔隙)多,而在降雨(入渗)初期土壤水处于不饱和状态,入渗率主要受土水势和土壤大孔隙流的控制[17]。这与武晓莉等[18]、胡阳等[19]研究结论相一致。武晓莉等[18]对晋西黄土区典型植被类型土壤水分特性研究发现,非毛管孔隙对土壤入渗的影响最大,其与初始入渗速率呈显著正相关关系。同样,胡阳等[18]对岩溶坡地荒地、草地、灌丛和林灌不同植被类型土壤水分入渗特征及影响因素的研究亦发现,影响土壤初始入渗速率、稳定入渗速率的主要因素为土壤非毛管孔隙度,且呈正相关关系。自然生生草区土壤自然含水率最低,而自然生草区土壤的初始入渗速率最高,这与郭晓霞等[20]研究结果一致,其对黄土高原土壤水分入渗特性进行研究发现,当0~20 cm土壤含水率较低时,土壤水分初始入渗速率较高。稳定入渗速率是评价土壤物理特性的一个重要指标,本试验结果表明,不同小区土壤稳定入渗率为清耕区>自然生草区>大豆区。而赵梦杰等[21]研究表明,植被覆盖对稳定入渗率的影响比较明显,植被覆盖度越高,稳渗率越大;何子淼等[22]对黄丘区野外草被坡面土壤入渗参数变化规律与模拟的研究也得出相似结论。清耕区植被覆盖度为0,而清耕区稳定入渗率大于自然生草区和大豆区,这是由于清耕区人为的耕作除草,土壤较为疏松,不仅总孔隙度大,大孔隙也相对较多。而大豆区初始入渗率和稳定入渗率均最小,这可能与大豆区土壤体积质量大,且大孔隙少有关。刘澄静等[23]研究亦表明,土壤渗透特性与土壤体积质量、总孔隙度存在显著相关关系,其中总孔隙度对土壤渗透性能产生的影响作用最大;陈家林等[8]对太行山低山丘陵区不同植被类型土壤渗透特性及影响因素的研究也得出相同结论。土壤体积质量越小,总孔隙度越大,土壤水分入渗速率越大,入渗速率与体积质量呈负相关,与总孔隙度呈正相关。不同植被覆盖区土壤入渗速率均表现为:初始入渗速率>平均入渗速率>稳定入渗速率,这与闫建梅等[17]研究川中丘陵区甜橙林、核桃林、水保林、梯平地、缓坡耕地这 5种不同土地利用方式土壤入渗特征的研究结果一致。
运用Philip、Horton、Kostiakov模型这3种经典模型对红枣经济林不同植被覆盖土壤水分入渗过程进行拟合发现,Horton入渗模型对红枣经济林不同植被覆盖区土壤入渗过程拟合效果最好,这与武晓莉等[18]对晋西黄土区的研究结论相似。而张志华等[25]对太行山低山丘陵区土壤渗透性进行研究则发现,Kostiakov拟合效果最好,Horton模型次之,Philip模型拟合效果最差。研究区域的不同可能是造成模型拟合效果差异的主要原因,但还需进一步深入研究
5 结 论
1)植被覆盖影响红枣经济林土壤水分入渗特征。清耕对照区土壤的初始入渗速率和稳定入渗速率分别为4.058 5 mm/min和1.952 8 mm/min,自然生草区明显提高了土壤初始入渗速率,达 4.902 0 mm/min;大豆区土壤初始入渗速率和稳定入渗速率均低于自然生草区和清耕区。
2)自然生草提高了植被盖度,还有效提高了土壤非毛管孔隙度,从而提高了初始入渗速率;与清耕相比,大豆区降低了土壤总孔隙度和非毛管孔隙度,降低了土壤初始入渗速率和稳定入渗速率。
3)Horton模型拟合效果最好,Philip模型次之,Kostiakov模型的拟合效果较差。
猜你喜欢
河北地质(2022年2期)2022-08-22 06:24:04
华南农业大学学报(2021年4期)2021-07-24 06:02:14
干旱地区农业研究(2019年3期)2019-07-18 07:09:40
现代园艺(2017年23期)2018-01-18 06:58:12
山东工业技术(2017年21期)2017-11-04 21:21:04
高原山地气象研究(2016年2期)2016-11-10 06:06:27
应用海洋学学报(2015年2期)2015-11-22 07:36:28
塔里木大学学报(2014年3期)2014-03-11 18:47:27
河南科技(2014年4期)2014-02-27 14:07:25
自然资源遥感(2014年2期)2014-02-27 11:56:12
