
迁西地区39份板栗种质叶片的功能性状多样性1)
2020-05-13 00:51刘亚斌郭素娟
东北林业大学学报 2020年4期
刘亚斌 郭素娟
(省部共建森林培育与保护教育部重点实验室(北京林业大学),北京,100083)
板栗(CastanetsmollisimaBlume)属于壳斗科(Fagaceae)栗属(Castanea)植物,是多年生落叶乔木。其营养含量十分丰富,含有较高的糖类和淀粉以及多种矿质元素等[1],价值极高,自古以来就有“树上粮食”,“铁杆庄稼”的美称[2-3],是我国的特产树种。与枣、桃、杏、李并列为我国古代5大名果[4]目前,我国发现并确定的板栗品种有300多个,建立4处栗属种质资源库,收集并保存6个种,347份种质材料[5]。根据产区的气候条件、土壤质地、栽培管理、人工定向选择以及品种性状特性等因素,分为6个地方品种群,即华北品种群、长江流域品种群、西北品种群、东南品种群、西南品种群和东北品种群[6]。
生物多样性是遗传多样性与环境多样性的综合体现,表现型和基因型之间存在着基因表达、个体发育、环境调控等复杂的中间环节,根据表现型差异来反映基因型差异具有重要的意义[7]。相关研究表明,植物叶片表型变异是遗传变异的重要特征之一[8]。叶片是植物连接自身与外界环境的重要纽带,是植物将自然界中的能量转换并自我利用的重要器官[9],树体及果实中大部分营养物质来源于叶片的光合作用。叶片的功能性状可直接影响植物的基本行为和其功能运作[10]。同一物种,不同群体的植物在适应同一环境过程中,其叶片功能性状存在差异[11]。
目前板栗生物多样性的研究主要以枝条、叶片、花序、果实为研究对象。李颖等[12]人以10个省份板栗群体,164份板栗种质为材料,通过对结果母枝9个表型性状进行一般分析、系统分组设计、聚类,结果表明:板栗结果母枝的9个表型性状在群体内和群体间变异丰富,聚类结果表明板栗群体表型特征基本按地理距离而聚类;江锡兵等[13]人以不同板栗种质资源的叶片为研究对象,以质量性状、数量性状来反映不同地理群体板栗叶片的表型多样性;江锡兵,龚榜初等[14]人对10个省份90个板栗种质的叶片及坚果的表型性状进行了多样性分析,以各个性状的平均变异系数来表明其遗传的稳定性,用表型分化系数来探讨其性状的主要变异来源等。
叶片功能性状可以很好的反映出植物对资源的获得、利用及利用效率,能够反映植物适应环境的生存对策[15]。虽然前人已经对不同板栗种质资源的叶片进行了研究,但是关于迁西地区不同产地来源的板栗叶片功能性状差异研究较少。迁西地区做为全国知名的板栗种植地区,了解不同板栗种质资源叶片功能性状在迁西地区所表现出的差异及其丰富的多样性,对于探讨板栗对迁西地区环境的适应能力、不同种质板栗叶片功能性状的综合评价及对迁西地区板栗的引种、选育新种质、新品种等工作具有重要意义。
1 材料与方法
以源自7个板栗主产省份的39份板栗种质资源为试验材料。根据地域分布,将39份板栗种质资源共划分为7个群体,即北京群体、河北群体、山东群体、贵州群体、湖北群体、安徽群体、江苏群体。
试验材料定植于河北省迁西县东寨板栗种质资源圃,在该试验地,南北方向每列种植34株板栗,株行距2 m×3 m,南北两侧种植两行保护行,每列包含有3个不同板栗种质资源,每个板栗种质资源定值10株,均为2013年以接穗的形式嫁接到3年生实生母树砧木上。同时,试验地气候、土壤等条件以及其它管理措施相一致。其种质编号、名称及来源如表1。

表1 板栗种质名称及其来源
2018年8月上旬(叶片成熟期)采集板栗叶片,在每个种质的10棵树中选择长势一致并能代表该种质的5株试验树,挂牌标记并记录,每株试验树选择树冠中部外围东、南、西、北4个方向的结果枝,在结果枝着生栗蓬或雌花簇的部位选择长势良好、无明显缺陷且无病虫害的5枚叶片,并挂牌标记。共计挂牌叶片数3 900枚。2018年9月(果实成熟期)采集果实,在每棵试验树树冠中部外围东、南、西、北4个方向分别采摘5个长势良好且无病虫害的刺苞,每个种质共采集100个刺苞。
叶片DSPA值测定:8月上旬(叶片成熟期),在晴天上午10:00—11:00进行试验[16]。采用便携式叶绿素仪(SPAD-502)测定不同种质板栗已挂牌叶片的DSPA值,以DSPA值来反映叶绿素相对含量。在每个叶片的叶脉旁测定叶片上中下3个部位的DSPA值,每个部位测定3次,共9次,取其平均值[17],并记录。
叶片表型性状测定:采集39组,共计3 900枚叶片。采集完毕后于冰箱4 ℃暂存,采用便携式叶面积仪进行叶片扫描,测定叶面积(AL)、叶周长(PL)、叶片横径(DL)、叶片纵径(DLV)、叶形指数(IL)[9]等指标。最后将叶片放入烘箱内60 ℃烘干至衡质量,记录叶片干质量(LDW),计算比叶质量[18](ACSL)。
叶形指数(IL)=叶片纵径(DLV)/叶片横径(DL)。
(1)

(2)
坚果单粒质量测定:2018年9月(果实成熟期)采集果实,每个种质共采集100个刺苞,将其去苞保留坚果,统计坚果粒数。单粒质量的测定:采用电子天平称取30粒坚果总质量,再计算出平均单粒质量,平行测定3次,取平均值[19]。
数据分析:利用Excel2010进行数据记录与制作表格,SPSS20软件进行双因素巢式设计方差分析,R语言软件数据进行相关性分析和聚类分析。
采用双因素巢式设计方差分析,线性模型为:
Yijk=μ+Si+T(i)j+ε(ij)k。
式中:Yijk为第i个群体第j个家系第k个观测值,μ为总均值,Si为群体效应(固定),T(i)j为群体内家系效应(随机),ε(ij)k为试验误差。
VST为表型分化系数,表示群体间变异占遗传总变异的百分比分比[12,14,20],式中:δ2t/s为群体间方差分量,δ2s为群体内方差分量。
VST(%)=[δ2t/s/(δ2t/s+δ2s)]。
(3)
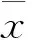
(4)
2 结果与分析
2.1 板栗叶片功能性状种质间多样性
叶片是植物进行光合作用的主要场所[9],与果实营养物质的积累具有密切的相关性[21],直接关系到植物的经济产量。叶片功能性状的多样性反映了植物对外界环境的适应能力以及对资源的利用效率[22]。通过对41份种质板栗叶片功能性状进行一般性分析,由表2可发现,板栗叶片8个功能性状在种质间具有极显著差异(P<0.01),表明不同来源的板栗叶片的功能性状在种质间存在广泛变异。其中叶面积(AL)均值的极大值与极小值分别是185.29和117.99 cm2,极差为67.30 cm2;叶周长(PL)均值的极大值与极小值分别是228.15和107.48 cm,极差为120.67 cm;叶片纵径(DLV)平均值的极大与极小值分别是23.37和19.84 cm,极差为3.53 cm;叶片横径(DL)平均值的极大与极小值分别是15.03和9.05 cm,极差为5.98 cm;叶形指数(IL)均值的极大值与极小值分别是2.61和1.59,极差为1.02;叶片干质量(LDW)均值的极大值与极小值分别是1.50和0.75 g,极差为0.75 g;比叶质量(ACSL)均值的极大值与极小值分别是9.96×10-3和4.68×10-3g/cm2,极差为5.28×10-3g/cm2;叶片DSPA均值的极大值与极小值分别是55.62和43.32,极差为12.30。

表2 不同板栗种质叶片部分功能性状比较

续(表2)

种源编号叶形指数(IL)叶片干质量(LDW)/g比叶质量(ACSL)DSPA值安徽392.29ABCDE 1.20ABCDEFG6.68CDEFGH51.09BCDE 北京252.08CDEFG0.80IJ6.82CDEFGH50.03BCDEFG261.97CDEFG0.92EFGHIJ6.92CDEFGH45.85FGH272.15BCDEFG0.99EFGHIJ6.47DEFGHI50.31BCDEF282.61A1.17BCDEFG8.36ABCD45.15GH291.89EFGH0.83HIJ5.44GHI50.69BCDEF302.35ABC0.93EFGHIJ7.55BCDEF49.05BCDEFGH311.97CDEFGH1.22ABCDEF6.85CDEFGH49.31BCDEFG贵州332.05CDEFG0.88FGHIJ6.19FGHI49.01BCDEFGH342.09CDEFG1.02DEFGHIJ7.55BCDEF51.06BCDE河北12.08CDEFG1.14BCDEFGI7.55BCDEF47.85CDEFGH22.04CDEFG0.85GHIJ6.37EFGHI49.47BCDEFG32.24ABCDEF0.94EFGHIJ6.72CDEFGH48.05CDEFGH42.05CDEFG1.07CDEFGHIJ6.79CDEFGH48.83BCDEFGH52.13BCDEFG1.43AB9.96A47.04DEFGH61.97CDEFGH0.75J4.68I44.32H72.33ABCD1.11BCDEFGI8.13BCDE50.65BCDEF82.18BCDEFG1.03DEFGHIJ6.98CDEFGH47.55DEFGH92.51AB1.40ABC8.53ABC55.62A102.22ABCDEF1.20ABCDEF7.48BCDEF50.49BCDEF112.10CDEFG1.15BCDEFGI8.10BCDE49.61BCDEFG122.37ABC1.50A8.86AB50.85BCDE132.08CDEFG1.13BCDEFGI6.93CDEFGH51.58ABCD142.09CDEFG1.05DEFGHIJ6.36EFGHI49.93BCDEFG152.06CDEFG1.26ABCDE8.13BCDE48.48CDEFGH161.90EFGH1.34ABCD7.16BCDEFG46.36EFGH172.27ABCDEF1.02DEFGHIJ8.31ABCD48.21CDEFGH182.21ABCDEF1.16BCDEFG7.48BCDEF46.53EFGH192.24ABCDEF1.03DEFGHIJ6.91CDEFGH50.94BCDE201.87FGH1.10BCDEFGI7.54BCDEF48.55CDEFGH212.07CDEFG0.88FGHIJ5.18HI47.57DEFGH221.91DEFGH0.89FGHIJ5.99FGHI48.70CDEFGH231.59H1.08CDEFGHIJ6.17FGHI53.69AB242.07CDEFG0.95EFGHIJ6.80CDEFGH47.23DEFGH湖北352.12BCDEFG0.93EFGHIJ5.70FGHI48.75CDEFGH362.13BCDEFG0.99EFGHIJ6.16FGHI52.71ABC372.23ABCDEF1.06CDEFGHIJ6.84CDEFGH50.93BCDE江苏381.80GH1.02DEFGHIJ6.59DEFGH47.91CDEFGH山东322.15BCDEFG1.10BCDEFGI7.40BCDEF50.57BCDEF
注:同列不同的大写字母表示为在0.01水平上的相关性显著。
2.2 板栗叶片功能性状群体间多样性
从表3可得到,在7个群体间,安徽板栗群体叶片表现为叶形较大,叶片干质量(LDW)及叶片DSPA值,在7个板栗群体中含量最高;江苏板栗群体在7个板栗群体间,具有较大的叶片横径(DL),较小的叶形指数(IL),均与与其他6个板栗群有显著的差异,北京、河北、山东在地理位置相互毗邻,遗传距离相近,其叶片的功能性状最为接近。

表3 不同板栗群体部分叶片功能性状均值及方差分析
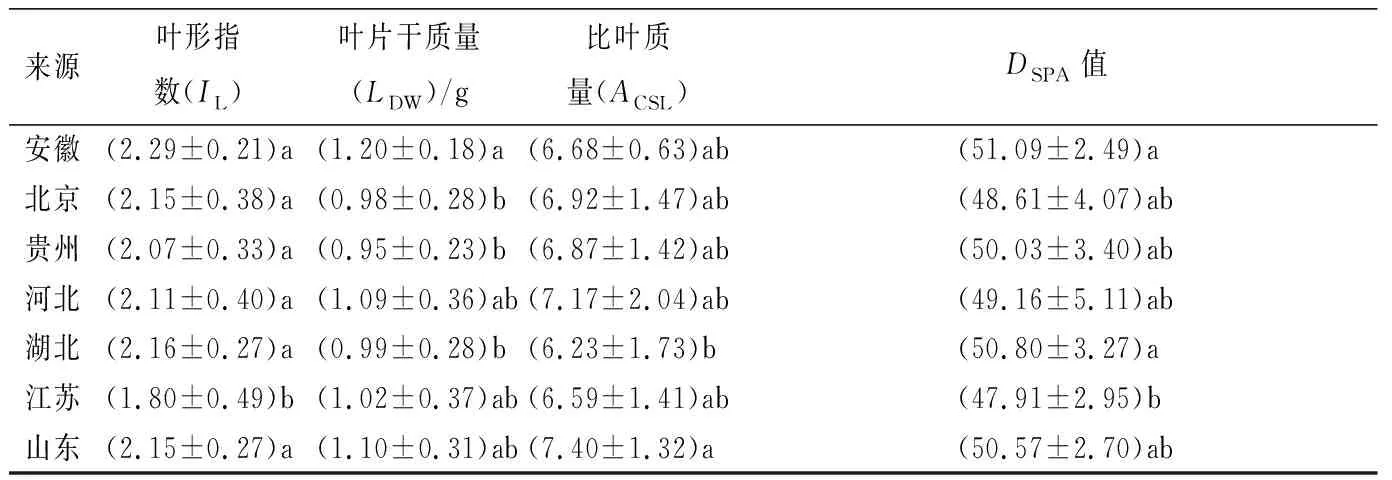
来源叶形指数(IL)叶片干质量(LDW)/g比叶质量(ACSL)DSPA值安徽(2.29±0.21)a(1.20±0.18)a(6.68±0.63)ab(51.09±2.49)a北京(2.15±0.38)a(0.98±0.28)b(6.92±1.47)ab(48.61±4.07)ab贵州(2.07±0.33)a(0.95±0.23)b(6.87±1.42)ab(50.03±3.40)ab河北(2.11±0.40)a(1.09±0.36)ab(7.17±2.04)ab(49.16±5.11)ab湖北(2.16±0.27)a(0.99±0.28)b(6.23±1.73)b(50.80±3.27)a江苏(1.80±0.49)b(1.02±0.37)ab(6.59±1.41)ab(47.91±2.95)b山东(2.15±0.27)a(1.10±0.31)ab(7.40±1.32)a(50.57±2.70)ab
注:同列不同的小写字母表示为在0.05水平上的相关性显著;*表示P<0.05,** 表示P<0.01;表中数据为“平均值±标准差”,平均值为总数据均值。
采用双因素巢式分组试验设计方差分析,探究不同来源板栗叶片功能性状在群体间和群体内两个层次上的差异显著性,如表4,结合表2、表3,板栗叶片的相关8个功能性状不仅在群体内差异得到了极显著水平,并且在群体间也达到了极显著的水平,说明板栗叶片各功能性状在群体间和群体内多样性丰富,均存在着广泛变异,具有较好的育种潜力。

表4 板栗叶片功能性状群体间和群体内的方差分析
注:*表示P<0.05差异显著;** 表示P<0.01差异极显著。
2.3 板栗叶片功能性状群体内多样性
由表5可得,板栗叶片8个功能性状指标的平均变异系数在6.91%~27.57%之间。其中叶片干质量的平均变异系数最大为27.57%,说明叶片干质量存在较大的群体差异,不同板栗群体间叶片干物质含量存在较大差异,具有较大的遗传变异潜力。
不同群体叶片功能性状变异系数均值存在较显著差异,表明各群体的叶片功能性状呈现多样性。板栗叶片功能性状不同群体间比较时,江苏板栗群体的各性状的平均变异系数均值最大,为23.30%,其次为河北板栗群体(22.05%),而安徽群体的各性状平均变异系数均值最小,为9.36%,表明该群体表型多态性与其它群体相比较为亏乏,稳定性较好。不同群体内性状多态性丰富程度由高到低依次为:江苏、河北、北京、贵州、湖北、山东、安徽。

表5 不同板栗群体叶片功能性状平均变异系数 %
2.4 板栗叶片功能性状多样性来源
按双因素巢式设计方差分量比组成,进一步分析出各方差分量占总变异的比例。由表6可得,方差分量百分比在群体间和群体内的范围分别是27.75%~55.88%和33.24%~62.25%。板栗叶片功能性状在群体遗传组成上存在较大差异;各性状表型分化系数(VST)存在较大差异,表型分化系数(VST)值变化范围为30.83%~62.70%,平均值为46.05%,即群体间的平均变异与群体内的平均变异比约为6∶7,表明各性状群体内变较群体间变异大,说明群体内变异是板栗叶片8个功能性状呈多样性的主要来源。
2.5 板栗叶片功能性状相关性
叶片是植物自身与外界环境进行物质交换的主要器官,通过利用水分、CO2、太阳能等进行自身物质的积累,与果实有机物的积累具有密切相关性[21],直接关系到植物的经济产量。叶片的功能性状可直接影响植物的基本行为和其功能的运作[10],同时能够反映植物遗传特性以及植物适应周围环境变化的响应对策[23]。
植物叶片的功能性状、化学组分和生理结构存在相互依存的内在联系,并且这种内在联系普遍存在于植物种群、群落和生物区系中[21]。对板栗叶片8个功能性状指标与坚果单粒质量进行相关性分析。由表7可得出,板栗叶片各个功能性状间存在一定的相关关系,既相互联系又相互独立。而DSPA值表示叶绿素的相对含量,是叶片生理功能性状,与叶片表性功能性状间没有表现出显著的相关性。坚果单粒质量与叶面积、叶片纵径呈显著正相关(P<0.05),与DSPA值呈极显著正相关(P<0.01),说明板栗叶片的受光面积与叶片DSPA值对板栗坚果的单粒质量有显著的影响。因此板栗叶片面积及DSPA值可以作为迁西地区引种及选育工作的一个重要参考指标。

表6 板栗群体叶片功能性状方差分量和群体间功能性状性状分化系数

表7 板栗叶片功能性状及单粒重综合相关性分析
注:*在0.05水平上的相关性显著,** 在0.01水平上的相关性极显著。
2.6 板栗叶片功能性状聚类分析
相关研究指出,根据产区的气候条件、土壤质地、栽培管理、人工定向选择以及品种性状特性等因素,分为6个地方品种[6]。在该研究中北京、河北、山东板栗群体属于华北板栗品种群,江苏、安徽、湖北板栗群体属于长江流域板栗品种群,贵州板栗群体属于西南板栗品种群。在采用系统聚类法并对7个板栗群体39份板栗种质资源进行聚类分析后发现:在欧氏距离为7时,7个板栗群体被分为2大类,即河北、北京、山东、贵州板栗群体聚为一类(Ⅰ类),江苏、安徽、湖北群体群体聚为一类(Ⅱ类),其中河北、北京、山东均位于北方,地理位置上相互临近,遗传距离相近,气候相似,因此其叶片功能性状相似,聚为一类。江苏、安徽、湖北均在长江中下游,根据上述理论,其板栗群体也聚为一类。但是,贵州板栗群体属于西南板栗品种群,与河北、北京、山东聚到Ⅰ类中,就参试板栗种质资源来,说结合表2、表3表明:河北、北京、山东、贵州板栗群体叶片功能性状接近。
3 结论与讨论
本研究中,板栗叶片8个功能性状指标在各个种质间,群体间与群体内均存在显著差异。在各个功能性状间,叶值DSPA值反映叶绿素的相对含量,是叶片生理功能性状,变异系数为6.91%,与板栗叶片表型功能性状(19.89%)相比,其变异系数较小,离散程度低。根据双因素巢式设计方差分量比组成,其表型分化系数为46.05%,群体间变异较群体内变异小,表明群体内变异是板栗叶片8个功能性状呈多样性的主要来源。板栗叶片各个功能性状间存在一定的相关关系,既相互联系又相互独立。研究结果表明板栗叶片的受光面积与叶绿素相对含量对板栗坚果的单粒质量有显著的影响。聚类分析中,将7个省份(群体)聚为2大类,河北、山东、北京、贵州板栗群体聚为一类,安徽、江苏、湖北板栗群体聚为一类,其聚类结果与地理分布格局大致吻合。
了解种质资源的遗传多样性是育种工作前提和基础,对挖掘有益种质、引种栽培、发现新种质等多方面具有重要意义[24]。本文结果显示,板栗叶片的8个功能性状在群体间和群体内均存在极显著差异(表1~2),这与李颖等人对10个板栗群体结果母枝相关表型性状的研究[12]及周连第等人对8个主产地板栗叶片表型性状多样性研究结果相似[25]。说明板栗叶片8个功能性状在群体间和群体内均存在丰富变异,多样性水平很高,育种选择潜力巨大。
变异系数(VC),又称“离散系数”,是概率分布离散程度的一个归一化量度,反映数据离散程度的绝对值。变异系数越大,则性状值离散程度越大[26],可用它来比较衡量不同的性状之间的变异程度,同时变异系数的大小也可也作为性状选择改良潜力的一个重要指标[12]。本研究中,同一群体不同性状间、同一性状不同群体间的变异程度均存在差异。表型形态性状的差异是基因与生态环境共同作用的结果,说明群体间板栗的基因型和环境异质性导致群体表型变异的差异,也体现了群体间板栗种质资源在育种上的不等值性。本研究中叶片功能性状平均变异系数的大小依次为叶片干质量、叶片周长、比叶质量、叶片横径、叶面积、叶形指数、叶片纵径、DSPA值,为选择具有观赏性的板栗种质资源提供了依据。;江苏、河北板栗群体的各性状的平均变异系数均值较大,性状遗传稳定性较低,表明该群体叶片功能性状多态性较为丰富。在育种工作中,考虑不同板栗群体平均变异系数大小的同时,还要关注群体内的极性个体。
生物群体间的变异反映了地理、生殖隔离上的差异,是生物多样性的重要组成部分[12],它在某种程度上反映了该生物对不同生存环境适应的广泛程度,生态位宽度的大小。其值越大适应的环境越广[27]。该研究的7个板栗群体共39份板栗种质资源,群体间8个功能性状的表型分化系数(VST)值介于6.91%~27.57%之间,平均为46.05%,群体间的平均变异与群体内的平均变异比约为6∶7,说明就板栗叶片8个功能性状,其变异的主要来源是群体内变异。此研究结果与江锡兵[14]基于板栗叶片、果实表型性状群体间、群体内的平均表型分化系数研究所提出的群体内变异远高于群体间变异的研究结果相似。
相关研究表明,植物叶片的功能性状、化学组分和生理结构存在相互依存的内在联系,并且这种内在联系普遍存在于植物种群、群落和生物区系中[21]。通过对7个群体,39份板栗种质资源叶片的8个功能性状指标与坚果单粒质量进行相关性分析。结果表明,板栗叶片的受光面积与叶片DSPA值对板栗坚果的单粒质量呈显著的正相关。说明在迁西地区叶面积大,叶绿素相对含量高的板栗种质,其板栗坚果单粒质量也越大。因此板栗叶片面积及叶片DSPA值可以作为迁西地区引种及选育工作的一个重要参考指标。
在该研究中河北、北京、山东板栗群体聚为一类,这3个省份在地理位置上相互毗邻,又同属于华北板栗品种群,其遗传距离相近,气候相似,因此,其叶片功能性状相似,聚为一类;江苏、安徽、湖北群体群体聚为一类,虽然这3个省份不处于同一纬度上,湖北与江苏相隔,但该3个板栗群体同属于长江流域板栗品种群,故聚为一类。聚类情况基本上按地理距离而聚类,揭示了群体间叶片功能性状变异的连续性[12]。
生物表型形态变异是由其遗传组成和生态环境的共同作用下产生的,是群体生物遗传信息的重要表征,也是长期适应与进化的产物。环境与栽培管理措施对植物表型性状的影响较大,在相同的生长环境、栽培管理等条件下,来探究植物不同性状的多样性能够更好的研究其遗传本质[28-30]。在该研究中的不足之处在于,所涉及到的试验材料来源于7个板栗主产省份,试验材料定植于河北省迁西县东寨板栗种质资源圃,虽然保证了其生长环境、栽培管理等条件的一致性,但未能探究板栗表型形态多样性与来源地生态因子间的相关性。同时,栗属植物为雌雄同株,其雌雄花序较为明显,为异交植物,同时其分布范围有较大的重叠,存在自然杂交的可能[31]。因此,在育种工作中应运用人工授粉、亲本回交等方法,在分子水平上利用不同类型的优良基因资源进行基因重组等,才能更加深入的揭示板栗的交配机制;在引种工作中该研究现有不足之处,虽然通过聚类结果可以考虑将同一类群的板栗群体进行相互间的引种,但在该研究中,未涉及到不同板栗种质资源在迁西地区的适应性以及产量等指标。因此这些问题均有待于进一步的探讨与研究。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
中国现代医生(2022年21期)2022-08-22
农村科学实验(2022年2期)2022-03-12
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
快乐作文(1.2年级)(2021年12期)2021-03-21
三农资讯半月报(2020年2期)2020-03-09
含笑花(2019年6期)2019-12-09
