
猴樟苗期生长规律及植株养分动态变化
2020-05-11 06:06于志民唐佳乐涂淑萍
南方农业学报 2020年3期
于志民 唐佳乐 涂淑萍


摘要:【目的】探讨猴樟苗期生长规律及其养分动态变化规律,为猴樟苗期培育合理水肥管理提供理论参考依据。【方法】测定猴樟一年生容器苗苗高和地径生长、生物量积累和分配及苗体氮、磷、钾含量的年变化,利用Logistic非线性模型对苗高和地径生长规律进行拟合与分析。【结果】猴樟一年生苗苗高有2次生长高峰,高峰期净生长量占全年净生长量的55.87%;地径有3次生长高峰,高峰期净生长量占全年净生长量的47.4%,地径生长高峰滞后于苗高生长高峰。9月为苗木生物量积累高峰期,6—8月中旬幼苗以地上部生物量积累为主,其生物量占整株比例从67.1%升至84.4%;根系生长于8月中旬以后开始加速,生物量占整株比例从15.6%升至20.2%。利用Logstic方程建立猴樟一年生苗高、地径生长规律的拟合模型,其决定系数(R2)分别达0.985和0.994,可靠性较高;将猴樟一年生容器苗生长划分为3个时期:渐增期、速生期、缓增期。生长初期,植株根、茎、叶的氮磷含量均有所增加,而钾含量表现为根部下降,茎叶上升;生长后期根部的磷钾含量上升,氮含量下降;氮元素主要分布于叶部,钾和磷元素主要分布于根茎部。【结论】猴樟一年生苗地径生长滞后于苗高生长,幼苗生长具有明显的阶段性,且苗木各器官氮、磷、钾含量随时间而发生动态变化;建议在猴樟苗期管理过程中氮、磷、钾应配合施用,生长后期应少施氮肥,增施磷、钾肥。
关键词: 猴樟;生长规律;养分动态;Logistic模型;水肥管理
0 引言
【研究意义】猴樟(Cinnamomum bodinieri Levl.)是樟科樟属乔木,原产于中国,多分布于云南、湖北、贵州、四川东部及湖南西部等地(杨成华和任远,2001),是我国亚热带常绿阔叶林中重要的组成树种之一(于志民等,2016)。猴樟具有生长速度快、树形美观、抗逆性强及木材紧密的特性,是优良的用材木(韩浩章等,2019);此外,其适宜生长于深厚、肥沃和水湿条件较好的土壤,对水分和温度具有广泛的适应性(韦小丽和熊忠华,2005),被广泛应用于园林绿化(于志民等,2017)。树木的生长发育不仅受到植物本身遗传物质的影响,还与各类生态因子息息相关,了解幼苗的生长规律不仅有利于优质苗木的培育,降低育苗成本,且通过结合植物生长过程中养分运转规律,对猴樟育苗过程中水肥管理制度的合理制定具有十分重要的意义。【前人研究进展】韦小丽(2003)对不同光环境下香樟与猴樟苗木的生态适应性进行了分析比较,发现猴樟比香樟稍喜光。程许娜(2012)、宋芳琳等(2012)通过自然降温试验发现猴樟较芳樟、香樟具有更强的低温胁迫适应能力,猴樟对-7 ℃能表现出一定的抗寒能力,但短期内温度骤降会对猴樟幼苗造成一定伤害。张怡等(2014)通过对云南及贵州地区6个地理种源猴樟进行育苗试验,初步筛选出云南腾冲地区种源猴樟为优良种源,贵州凯里地区猴樟对光的生态适应性较好。韦小丽和陈瀚林(2016)对猴樟混交林进行研究发现猴樟×杜英×楠木是一种较好的混种模式。于志民等(2016,2017)对猴樟容器育苗展开研究,探索出最适宜猴樟一年生容器苗的育苗容器及育苗基质。杨应(2018)在研究中发现高浓度的CO2与AM真菌对猴樟幼苗生长和养分利用可能存在一定的协同作用。此外,贡丹敏等(2019)研究发现猴樟幼苗具有一定的耐盐碱胁迫能力;韩浩章等(2019)研究发现高浓度HCO3-和CO32-会降低猴樟对氮素、磷素及铁素的吸收;张丽华和韩浩章(2019)研究发现2%浓度的木醋液能改变猴樟幼苗土壤养分状况。【本研究切入点】目前,有关猴樟幼苗在生长过程中对养分需求及运转规律的研究鲜有报道。【拟解决的关键问题】通过对猴樟一年生苗的年生长规律及植株不同器官的氮、磷、钾养分含量变化规律进行研究,为猴樟育苗过程中确定合理水肥管理制度提供理论依据。
1 材料与方法
1. 1 试验地概况
试验地设在江西省南昌市昌北经济技术开发区江西农业大学林学院花卉盆景基地,位于东经115°49′46.72″、北纬28°45′34.04″,属亚热带季风湿润气候,日照充足,1月平均气温5.7 ℃、最低气温-3.9 ℃,7月平均气温28.6 ℃、最高气温38.7 ℃,年降水量1600~1700 mm,年平均相对湿度78.5%。
1. 2 试验材料
试验树种为由高安市邓志平园林苗木公司提供的生长健壮、无病虫害、大小一致的一年生猴樟播种苗。育苗基质按体积比为黄心土∶火烧土∶草炭土∶河砂∶生物有机肥=5∶2∶1∶2∶1。基质理化性质为:pH 5.92,容重1.29 g/cm3,饱和持水率43.88%,总孔隙度62.26%,有机质含量49.9 g/kg,有效磷含量54.44 mg/kg,碱解氮含量72.24 mg/kg,速效钾含量433.21 mg/kg。育苗容器采用规格为15 cm×18 cm的白色无纺布容器袋。
1. 3 试验方法
于2016年6月10日将猴樟容器苗(平均苗高7.45 cm、平均地径1.88 mm)摆放在苗床上,苗床上覆盖遮光度为30%的遮阳网,地上铺设地膜,以防止幼苗根系扎入土中。种植后于每天傍晚6:00进行浇水养护。于6月17日—9月17日,设置3个重复,每重复随机选取10株苗木作为固定观测株,每10 d用钢卷尺测量苗高,用电子数显游标卡尺测定地径,并在苗床内随机选取10株平均株(苗高、地径尽可能接近上一次测量的固定观测株的苗高、地径平均值),将苗木掘起,将叶、茎、根3部分分别置于105 ℃干燥箱中杀青15 min,之后在65 ℃条件下烘干,分别测定叶、茎、根干重。烘干后的根、茎、叶粉碎后采用H2SO4-H2O2消煮法分別测定其氮、磷、钾含量(鲍士旦,2000)。植株不同器官氮含量测定采用开氏定氮法;各器官磷含量测定采用钒钼黄比色法;各器官钾含量测定采用火焰光度计法。
1. 4 统计分析
试验数据采用Excel 2003及Sigmaplat 12.5进行整理及绘图;使用SPSS 22.0的Logistic回归分析拟合苗木的生长规律。
2 结果与分析
2. 1 猴樟一年生苗苗高年生长规律
由图1可看出,猴樟一年生苗的苗高生长遵循“慢—快—慢”的趋势。在全年的生长过程中,苗高出现2次生长高峰,第1次生长高峰出现在8月中旬,苗高旬净生长量达10.62 cm,占全年苗高总生长量的33.07%,接下来的8月下旬,由于气温持续较高,天气干燥,水分不充足,苗高生长速率减缓;9月上旬苗高出现第2次生长高峰,但持续时间较短,苗高旬净生长量为7.32 cm,占全年苗高总生长量的22.80%。2次生长高峰期苗高累计净生长量占全年净生长量的55.87%。
2. 2 猴樟一年生苗地径年生长规律
由图2可看出,猴樟一年生苗地径生长量呈逐渐增加趋势。在全年生长过程中地径出现3次生长高峰,第1次出现在7月上旬,地径旬净生长量为0.836 mm,占全年地径总生长量的15.8%,7月中旬地径生长速率减缓;7月下旬地径生长出现第2次生长高峰,旬净生长量为0.686 mm,占全年总生长量的12.9%;9月上旬出现第3次生长高峰,旬净生长量为0.995 mm,占全年总生长量的18.7%。3次生长高峰地径累积生长量占全年生长量的47.4%。
2. 3 猴樟一年生苗的根、茎、叶生物量年变化规律
由表1可知,在整个生长季中,猴樟一年生苗的根、茎、叶生物量呈逐渐上升趋势。其中,单株根干重在观测期内增加3.069 g,茎干重增加4.401 g,叶干重增加7.935 g。由此可看出,在整个观测期内猴樟一年生幼苗叶干重增长量最大,其次为茎,说明幼苗地上部生物量积累大于根系生物量积累。
由图3可知,猴樟一年生苗生物量在根、茎、叶分配的不同时间段表现出不同特征。根生物量对整株生物量的贡献率呈现先下降后上升的变化趋势,由6月中旬的32.9%下降至8月中旬的15.6%,再上升至9月中旬的20.2%;茎生物量在整株生物量所占比重在前期(6月中旬—7月下旬)变化不大,7月下旬以后所占比重迅速增加,由7月下旬的12.7%上升到9月上旬的28.5%;叶生物量对整株生物量的贡献率呈现先增后减的变化趋势,由6月中旬的56.1%上升到7月下旬的69.5%,再下降至9月中旬的51.6%。可见地上部生物量6—8月中旬占整株比例从67.1%升至84.4%;总体而言,叶生物量对整株生物量的贡献率最高。
2. 4 猴樟一年生苗苗高、地径日生长量拟合与生长阶段划分
2. 4. 1 幼苗苗高Logistic模型的建立与拟合 猴樟一年生苗苗高拟合后参数如表2所示,猴樟一年生苗苗高Logistic方程拟合模型拟合度(R2)为0.985,因此,此方程具有较高的可靠性。
2. 4. 2 猴樟一年生苗苗高生长阶段划分 猴樟一年生苗苗高的Logistic曲线方程为:Y=61.053/(1+e2.610-0.035t)(R2=0.985,P<0.01),通过对方程进行求导,计算得出猴樟一年生苗苗高拟合方程的各项参数指标(表3),猴樟一年生苗理论极限生长量为61.053 cm,最高速生长点为(87,30.53),即苗木生长到第87 d(9月11日)日生长量达到最高值(30.53 cm),此时生长速率最高,为0.46 cm/d。此外,猴樟一年生苗苗高生长速率的拐点出现在第43 d(7月29日)和第131 d(10月25日),而该段时间的线性增长率为0.41 cm/d,线性增长量为38.37 cm,占其理论总生长量的62.85%。因此,根据拟合分析及幼苗地上部分实际生长情况,可将猴樟一年生苗的苗高生长阶段划分为3个阶段:6月17日—7月29日为苗木生长初期(渐增期);7月29日—10月25日共88 d,为苗木生长速生期;10月25日开始往后为苗木生长缓增期。
2. 4. 3 幼苗地径Logistic模型的建立与拟合 猴樟一年生苗地径拟合后参数如表4所示,Logistic方程拟合模型R2为0.994,因此,此方程具备较高的可靠性。
2. 4. 4 猴樟一年生苗地径生长阶段划分 猴樟一年生苗地径的Logistic曲线方程为:Y=16.566/(1+e2.253-0.019t)(R2=0.994,P<0.01),通过对方程进行求导,计算得出猴樟一年生苗地径拟合方程的各项参数指标(表3),猴樟一年生苗地径理论极限生长量为16.566 cm,最高速生长点为(112,8.29),即苗木生长到第112 d(10月6日)的日生长量达到最高值(8.29 cm),此时生长速率最高,为0.08 cm/d。此外,猴樟一年生苗地径生长速率的拐点出现在第47 d(8月2日)和第178 d(12月11日),而猴樟一年生苗在该段时间的线性增长率为0.07 cm/d,线性增长量为9.17 cm,占其理论总生长量的55.35%。因此,可将猴樟一年生苗的地径生长阶段划分为3个阶段:6月17日—8月2日为苗木生长初期(渐增期);8月2日—12月11日共130 d,为苗木生长速生期;12月11日开始往后为苗木生长缓增期。
2. 5 猴樟一年生苗根、茎、叶的氮、磷、钾含量年变化规律
2. 5. 1 根、莖、叶的氮含量年变化规律 由图4可知,猴樟一年生苗各器官氮含量存在较大差异,叶片氮含量最高,根系次之,茎的氮含量最低。从各器官氮含量年变化规律看:(1)根系氮含量全年出现3个谷值及3个波动区段,3个谷值对应的日期分别为6月下旬、7月下旬和9月上旬,3个波动区段分别是6月中旬—7月中旬、7月中旬—8月上旬及8月上旬—9月中旬;前2个区段变化幅度不大,第1区段根部氮含量由6月中旬的0.96%下降至6月下旬的0.88%,而后上升至7月中旬的0.99%,第2区段根部氮含量由7月中旬的0.99%下降至7月下旬的0.85%,而后上升至8月上旬的0.92%,第3区段变化幅度较大,氮含量由0.92%下降至9月上旬的0.56%后上升至9月中旬的0.88%。(2)茎氮含量全年均呈下降趋势,6月中旬—8月中旬期间下降趋势极缓,由0.62%下降至0.56%;8月中旬—9月中旬下降幅度加大,由0.56%下降至0.32%。(3)叶片氮含量于6月中旬—7月上旬上升,7月上旬—8月上旬下降,8月上旬—8月中旬上升,但变化幅度均较小,约为0.10%;8月中旬后叶片氮含量大幅下降,至9月中旬,下降0.34%。可见,苗木生长期内氮主要分布在叶部,生长后期(9月中旬后)苗木地上部生长几乎停止,氮则向根部转移,有利于提高苗木的抗性,安全越冬;茎氮含量始终处于较低水平。
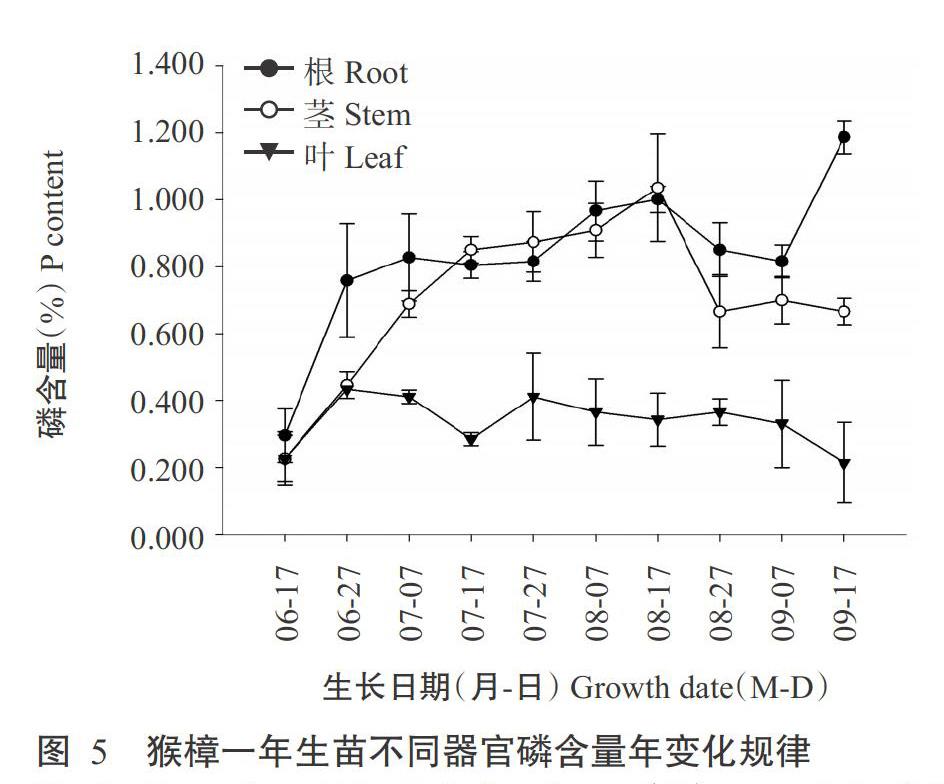
2. 5. 2 根、茎、叶的磷含量年变化规律 由图5可知,猴樟一年生苗根磷含量年变化呈现先增后减再增的变化趋势。生长初期的6月中旬—6月下旬和生长末期的9月上旬—9月中旬为根系磷快速增长期,分别由0.295%、0.816%增长至0.785%、1.187%;6月下旬—9月上旬根系磷含量先缓慢增加后缓慢下降,先增长到8月中旬的1.001%,再下降至9月上旬的0.816%,变化幅度很小。茎磷含量于定植初期的6月中旬—7月中旬增长迅速,随后增长缓慢,至8月中下旬,茎磷含量迅速下降。叶片磷含量较低,全年变化不大,生长初期(6月中旬)磷元素在根、茎、叶分配比例几乎相同,1个月后,根系和茎的磷含量高于叶片磷含量,至9月中旬地上部生长停止,磷向根系转移。因此,就全株磷含量而言,植株各器官磷含量表现为根系>茎>叶片。
2. 5. 3 根、茎、叶的钾含量年变化规律 由图6可知,根系钾含量总体呈下降趋势,且有3个谷值,分别出现在6月下旬、7月下旬和9月上旬,6月中下旬、7月中下旬和8月上旬—9月上旬钾含量均下降,生长后期(9月上中旬)根系钾含量增加。茎和叶片的钾含量年变化趋势大体相同,总体呈先增后减的变化趋势,生长初期钾含量增加,生长后期钾含量减少。因此,苗木旺盛生长期根系钾元素逐渐向茎和叶片转移,生长后期地上部生长停止后钾向根系转移。
3 讨论
本研究结果表明,猴樟一年生苗苗高及地径的生长遵循生物学的基本规律,表现为“慢—快—慢”的生长趋势,利用Logistic非线性方程对猴樟一年生苗苗高和地径生长曲线进行拟合具有较高的可靠性,根据Logistic非线性方程并结合苗木生长实际情况,将猴樟一年生苗年生长划分成3个时期,即渐增期、速增期、缓增期,其中苗高生长的速生期为7月29日—10月25日,持续88 d左右;地径的速生期为8月2日—12月11日,持续时间达130 d左右,由此表明猴樟一年生苗年生长具有明显的阶段性;但也有学者研究发现不同地区猴樟生长阶段存在一定的差异,贵州及云南地区种源的猴樟苗高及地径速生期在63~84 d(张怡等,2014),说明猴樟生长阶段可能与种源及气候环境有一定相关性。
不同生长阶段对苗木的管理方式、管理水平应有所不同。有研究表明,苗木生长差异主要来自于速生期的长短,速生期内苗木生长量约占整个生长期生长量的60%(程龙霞等,2015)。本研究中猴樟一年生幼苗苗高及地径在速生期内生长量分别占整个生长期生长量的62.85%和55.35%,可看出速生期对苗木生长具有极其重要的作用,其中速生期的长短对苗木当年生长量具有决定性作用,如贵阳市花溪区种源的猴樟幼苗苗高及地径在速生期内生长量分别占其整个生长期生长量的68.30%和43.20%(韦小丽和熊忠华,2005),两地猴樟幼苗在速生期内苗高生长量占比基本持平,而地径在速生期内生长量占比存在一定差异,究其原因,可能是因为两地猴樟幼苗速生期持续时间存在差异,本研究中猴樟地径速生期达130 d,而贵阳市花溪区种源的猴樟幼苗地径速生期仅持续62 d。因此,在苗木培育过程中,应加强速生期水肥管理,为苗木生长提供适宜的生长条件(吴国欣等,2010)。
本研究中,猴樟一年生苗地径生长滞后于苗高生长20 d左右。苗高于6月中旬开始生长,而地径从6月下旬开始增粗,生长起点滞后于苗高生长起点;地径于8月底达到生长拐点(生长速率最大),而苗高于8月上旬达到生长拐点;在苗高停止生长后20 d左右,地径停止生长。由此可见,在猴樟一年生苗生长年变化过程中,首先进行伸长生长,即苗高增加,在苗高生长了一段时间后再进行增粗生长,即地径的加粗,这与细胞分裂与伸长规律相似。在苗高生长初期,地径尚未生长的时期,苗木抗性很弱,尤其应注意病虫害防治,以及防风防雨等工作。
猴樟一年生苗氮含量分布为叶>根>茎,磷含量分布为根≈茎>叶,钾含量分布为生长初期(6月中旬)钾元素主要分布于根系,茎和叶片的钾含量较低,旺盛生长期根系钾逐渐向茎和叶片转移,生长后期地上部生长停止后钾向根系转移,这可能因为在猴樟一年生苗速生期其地上部生长旺盛,而此时正处于夏季,叶片蒸腾作用旺盛,K+大量向叶片转移参与气孔开合的调控,而地上部生长停止后根系仍处于生长阶段,K+向根系转移,参与矿质元素的跨膜运输。总体而言,苗木速生期应多施氮肥,促进地上部器官的生长,如苗高增长、地径增粗、叶片数量增加及地上部分干物质积累等;氮肥充足才能保证营养体正常生长,呈现出枝繁叶茂、树势强健的良好状态;而生长后期应少施氮肥,增施磷钾肥,以抑制地上部分的生长,防止地上部器官过于繁茂而消耗更多的有机物质,同时促进地下部根系生长,加速苗木硬化,从而提高苗木的抗性,确保其安全越冬。
4 結论
猴樟一年生苗地径生长滞后于苗高生长,幼苗生长具有明显的阶段性,且苗木各器官氮、磷、钾含量随时间而发生动态变化;建议在猴樟苗期管理过程中氮、磷、钾配合施用,生长后期应少施氮肥,增施磷、钾肥。
参考文献:
鲍士旦. 2000. 土壤农化分析[M]. 第3版. 北京:中国农业出版社:39-114. [Bao S D. 2000. Soil agrochemical analysis[M]. The 3rd Edition. Beijing: China Agriculture Press: 39-114.]
程龙霞,施曼,祝遵凌. 2015. 欧洲鹅耳枥一年生播种苗年生长动态探究[J]. 中国野生植物资源,34(5): 46-50. [Cheng L X,Shi M,Zhu Z L. 2015. Research on the dynamic growth of one-year-old Carpinus betulus[J]. Chinese Wild Plant Resources,34(5): 46-50.]
程许娜. 2012. 猴樟对低温胁迫的生理响应及其耐寒性分析[D]. 郑州: 河南农业大学. [Cheng X N. 2012. Physiological responses of Cinnamomum bodinieri Levl to low temperatrure and its cold tolerance[D]. Zhengzhou: Henan Agricultural University.]
贡丹敏,冒国娇,刘晓艳,马刘娟,殷瑜佳,顾吉,韩浩章. 2019. 苏打盐碱胁迫对猴樟幼苗渗透调节的影响研究[J]. 安徽农学通报,25(7): 23-25. [Gong D M,Mao G J,Liu X Y,Ma L J,Yin Y J,Gu J,Han H Z. 2019. Effect of soda salt and alkali stress on osmotic regulation of Cinnamomum bodinieri seedlings[J]. Anhui Agricultural Science Bulletin,25(7): 23-25.]
韩浩章,陈柳,李素华,张丽华,王晓立,张颖,王芳,蒋亚华. 2019. Na2CO3和NaHCO3混合处理对猴樟幼苗养分吸收的影响[J]. 东北林业大学学报,47(8): 8-13. [Han H Z,Chen L,Li S H,Zhang L H,Wang X L,Zhang Y,Wang F,Jiang Y H. 2019. Effect of Na2CO3 and NaHCO3 mixture on nutrient absorption of Cinnamomum bodinieri seedlings[J]. Journal of Northeast Forestry University,47(8): 8-13.]
宋芳琳,张苗苗,苏金乐,柴军舰,张广信. 2012. 自然降温过程中猴樟和香樟幼苗的抗寒性比较[J]. 西部林业科学,41(6): 48-52. [Song F L,Zhang M M,Su J L,Chai J J,Zhang G X. 2012. The comparison on cold resistance between Cinnamomum bodinieri and Cinnamomum camphora seedlings in natural decreasing process of air temperature[J]. Journal of West China Forestry Science,41(6): 48-52.]
韦小丽. 2003. 不同光环境下香樟、猴樟苗木的生态适应[J]. 山地农业生物学,22(3):208-213. [Wei X L. 2003. Ecological adaption of Cinnamomum camphora and Cinnamomum bodinieri seedlings in different light environment[J]. Journal of Mountain Agriculture and Biology,22(3): 208-213.]
韦小丽,陈瀚林. 2016. 不同类型猴樟混交林种间关系和冠层结构[J]. 山地农业生物学报,35(6): 1-8. [Wei X L,Chen H L. 2016. Theintraspecific relationship and canopy structure of different type Cinnamomum bodinieri mixed stand[J]. Journal of Mountain Agriculture and Biology,35(6): 1-8.]
韦小丽,熊忠华. 2005. 香樟和猴樟1年生播种苗的生长发育规律[J]. 山地农业生物学报,24(3):205-208. [Wei X L,Xiong Z H. 2005. Study on the growth pattern of Cinnamomum camphora Presl and Cinnamomum bodinieri Levl annual seedling[J]. Journal of Mountain Agriculture and Biology,24(3): 205-208.]
吴国欣,王凌晖,俞建妹,刘运华,张明慧,梁惠萍. 2010. 降香黄檀幼苗年生长节律研究[J]. 浙江林业科技,30(3): 56-60. [Wu G X,Wang L H,Yu J M,Liu Y H,Zhang M H,Liang H P. 2010. Study on growth rhythm of Dalbergia odorifera seedlings[J]. Journal of Zhejiang Forestry Science and Technology,30(3): 56-60.]
楊成华,任远. 2001. 优良园林树种猴樟及其培育[J]. 贵州林业科技,29(1): 28-31. [Yang C H,Ren Y. 2001. Elite tree species for landscape-Cinnamomum bodinieri and its cultivation[J]. Guizhou Forestry Science and Technology,29(1): 28-31.]
杨应. 2018. 提高CO2浓度下接种AM真菌对猴樟幼苗生长及养分利用的影响[D]. 贵阳:贵州大学. [Yang Y. 2018. Effects of AM fungi on the growth and nutrient utilization of Cirtnamomum bodinieri seedlings under increasing CO2 concentration[D]. Guiyang: Guizhou University.]
于志民,刘玮,邓志平,涂淑萍,邓光华. 2016. 不同类型及规格育苗容器对猴樟1年生苗生长的影响[J]. 南方农业学报,47(12): 2118-2123. [Yu Z M,Liu W,Deng Z P,Tu S P,Deng G H. 2016. Effect of container type and size on the growth of seedlings of Cinnamomum bodinieri Levl. annual seedlings[J]. Journal of Southern Agriculture,47(12): 2118-2123.]
于志民,涂淑萍,鄧光华,邓志平,刘玮. 2017. 不同配比基质对猴樟容器苗生长的影响[J]. 南方农业学报,48(12): 2218-2222. [Yu Z M,Tu S P,Deng G H,Deng Z P,Liu W. 2017. Effects of substrates with different ratios on seedling growth of Cinnamomum bodinieri Levl. container seedlings[J]. Journal of Southern Agriculture,48(12): 2218-2222.]
张丽华,韩浩章. 2019. 木醋液对猴樟幼苗土壤养分有效性的影响[J]. 西南林业大学学报,39(3):93-100. [Zhang L H,Han H Z. 2019. Effect of wood vinegar on soil nutrient availability of Cinnamomum bodinieri seedlings[J]. Journal of Southwest Forestry University,39(3): 93-100.]
张怡,韦小丽,王娈,陈模芳. 2014. 不同地理种源猴樟苗期生长变异性[J]. 西南农业学报,27(5):2162-2167. [Zhang Y,Wei X L,Wang L,Chen M F. 2014. Growth variability of Cinnamomum bodinieri seedlings from different geogra-phical provenances[J]. Southwest China Journal of Agricultural Sciences,27(5): 2162-2167.]
(责任编辑 邓慧灵)
