
云南程海和阳宗海季节性分层及其消退对冬季水华的潜在影响*
2020-05-08 02:39周起超王玮璐黄立成
湖泊科学 2020年3期
周起超 ,杨 炫,王玮璐,董 静,黄立成,秦 江
(1:云南大学生态与环境学院高原湖泊生态与治理研究院,昆明 650500) (2:云南省生态环境科学研究院云南省高原湖泊流域污染过程与管理重点实验室,昆明 650034) (3:河南师范大学水产学院,新乡 453007)
热力分层及其季节性变化是深水湖泊(水库)的一个重要特征,湖泊热力分层及其循环周期与稳定性对水体溶解氧(DO)[1]、沉积物微生物过程[2]、营养盐分布及其可利用性[3-4],以及浮游植物垂直分布与群落结构[3,5-6]等有着重要的影响,进而影响湖泊水质、初级生产力、食物网结构等,故对热力分层特征的深入了解是实现对深水湖泊生态环境保护的重要前提. 目前,国内外关于湖库热力分层的研究已有诸多报道,如针对Lake Müggelsee[7]、Lake Caldonazzo[8]、千岛湖[9-10]、天山天池[11]、纳木错[12]、天目湖[13]及空间大尺度的研究[14-15]等. 云南高原是我国云贵高原湖区的重要组成部分,由于缺乏相关基础性工作和基础数据的积累,目前关于其湖泊热力分层的研究仅局限于对抚仙湖[16-17]、泸沽湖[18-19]、洱海[20]、程海[21]和阳宗海[22]等的零星报道.
藻类(浮游植物)水华是诸多水体面临的严峻环境问题,表层水温常是浮游植物生物量季节变化(夏秋季高、冬春季低)的主控因素之一[23];稳定的热力分层被认为阻碍了水体上下层的物质交换,有助于藻类水华的暴发和维持[24-25],如气候变暖导致的更强烈的热力分层被认为利于夏季温跃层附近粉红浮丝藻(Planktothrixrubescens)水华的发生[26],人工加深温跃层深度亦有利于湖上层的P.rubescens占优[3],针对天目湖的研究显示,稳定的热力分层利于蓝藻门的繁殖[13]. 然而,浮游植物生物量的季节变化并不一定呈现“夏秋季高、冬春季低”的趋势,如云南程海浮游植物细胞密度、叶绿素a(Chl.a)浓度和初级生产力均会在冬季出现另一次波峰[27-28],根据文献记载[29]及笔者2015年1月19日的现场观测均显示程海在冬季发生过藻类水华(冬季水华,本研究以浮游植物生物量——Chl.a浓度间接表征),针对云南阳宗海的调查亦发现其冬季的Chl.a浓度明显高于夏季[30],但关于其发生机制仍不清楚.
因此,本研究基于2014年10月-2016年7月对程海和阳宗海开展4个季度的调查,在识别两个湖泊热力分层、垂直剖面DO与电导率(SpCond)分布及水柱表层营养盐、浮游植物生物量季节变化及其影响因子的基础上,重点探讨了两湖水体分层消退对冬季水华发生的潜在影响.
1 材料与方法
1.1 研究区域概况
程海(26°27′~26°38′N,100°38′~100°41′E;图1a)地处云南省丽江市永胜县中部、金沙江河谷地带,约1690年前后开始演变为内陆封闭型深水湖泊,水位1503.0 m,面积77.22 km2,平均水深25.7 m,最大水深35.1 m,蓄水量19.87×108m3,流域面积228.9 km2. 阳宗海(24°51′~24°58′N,102°58′~103°01′E;图1b)地处云南省昆明市东南,水位1770.75 m,面积31.68 km2,平均水深19.5 m,最大水深30.0 m,蓄水量6.17×108m3,流域面积192.0 km2. 程海和阳宗海均为云南九大高原湖泊之一、属构造断陷型中营养深水湖泊,为亚热带季风气候.
1.2 样品采集与指标测定
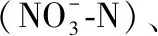
1.3 统计分析
本研究所示的均值为平均值±标准差;两独立样本(Mann-Whitney U)、多独立样本(Kruskal-Wallis H)非参数检验与Spearman相关性分析均由SPSS Statistics 24.0软件完成,P<0.05表示显著,P<0.01表示极显著.

图1 程海(a)和阳宗海(b)采样点位Fig.1 Sampling sites of Lake Chenghai(a) and Lake Yangzonghai(b)
2 结果
2.1 两湖气温与降水量的季节变化
根据国家气象信息中心(http://data.cma.cn/)资料(1981年1月-2010年12月),程海附近永胜县(台站编号:56652;经纬度:26.41°N,100.45°E;海拔:2130.5 m)1月、4月、7月、10月的月平均气温分别为6.4、14.9、19.1和14.5℃,月平均降水量分别为5.8、14.3、254和73.7 mm;宾川县(台站编号:56752;经纬度:25.5°N,100.34°E;海拔:1438.7 m)1月、4月、7月、10月的月平均气温分别为10.5、20.2、23.9和19.3℃,月平均降水量分别为3.4、9.4、139.8和50.1 mm. 阳宗海附近宜良县(台站编号:56880;经纬度:24.55°N,103.1°E;海拔:1532.5 m)1月、4月、7月、10月的月平均气温分别为9.1、19.2、21.6和17.2℃,月平均降水量分别为16.7、24.3、166.8和76 mm;澄江县(台站编号:56873;经纬度:24.41°N,102.55°E;海拔:1796.3 m)1月、4月、7月、10月的月平均气温分别为9.4、17.8、20.8和16.6℃,月平均降水量分别为14.2、27.6、210.5和80.6 mm.
2.2 两湖水温剖面与温跃层参数的季节变化
程海秋、冬、春、夏四季的表层水温分别为22.96±0.25、14.33±0.12、16.70±0.23和24.67±0.38℃(图2a~d),阳宗海秋、冬、春、夏四季的表层水温分别为19.98±0.17、13.63±0.16、18.01±0.69和23.43±0.33℃(图2e~h),两湖间四季或分层季节的表层水温均无显著性差异(P>0.05);除冬季外,两个湖泊秋、春、夏三季均有一定的热力分层,同一季节各点位的温跃层分布状况具有一定的异质性(图2,表1). 就温跃层的深度、厚度和强度而言(表1),程海春季温跃层深度显著大于夏季(P<0.01)和秋季(P<0.05),夏、秋季温跃层的深度、厚度和强度均无显著性差异(P>0.05),秋季的厚度和强度显著大于春季(P<0.01),夏季的厚度略大于春季(P=0.07)、强度显著大于春季(P<0.01);阳宗海夏季温跃层深度显著小于春季(P<0.01)和秋季(P<0.05)、春秋季间无显著性差异(P>0.05),秋季和春季的温跃层厚度和强度均无显著性差异(P>0.05),夏季的厚度显著大于秋季(P<0.05)和春季(P<0.01),夏季的强度与春季、秋季均无显著性差异(P>0.05). 就程海和阳宗海间温跃层参数比较而言(表1),程海的温跃层深度(P<0.01)、底界深度(P<0.05)均显著大于阳宗海,两湖温跃层厚度和强度则无显著差异(P>0.05).

图2 程海(a~d)和阳宗海(e~h)水温剖面的季节变化Fig.2 Seasonality in vertical profile of water temperature in Lake Chenghai (a-d) and Lake Yangzonghai (e-h)

表1 程海和阳宗海温跃层参数的季节变化
aN=5,bN=6,cN=8,dN=19,eN=3,fN=15.
进一步分析发现,程海表层水温与温跃层深度呈显著负相关(r=-0.654,P<0.01,N=19),与厚度(r=0.493,P<0.05,N=19)、强度(r=0.602,P<0.01,N=19)呈显著正相关;阳宗海表层水温与温跃层深度呈显著负相关(r=-0.536,P<0.05,N=15),与厚度呈显著正相关(r=0.568,P<0.05,N=15),与温跃层强度无显著相关性(r=0.246,P=0.376,N=15). 真光层深度与温跃层参数的关系方面,程海(N=19)或阳宗海(N=15)的真光层深度与温跃层深度、厚度和强度之间均未检出显著的相关性. 此外,程海和阳宗海的平均真光层深度分别为6.16±1.12 m(N=36)和6.44±1.21 m(N=24),两湖间四季或分层季节的真光层深度均无显著性差异(P>0.05).
2.3 两湖溶解氧浓度和电导率垂直分布的季节变化
程海DO浓度垂直分布的季节变化如图3a~d所示:在秋、夏季形成氧跃层,深水区达到缺氧状态(< 3 mg/L),秋季深层水DO浓度甚至低于1 mg/L;春季,形成氧跃层雏形,深水区暂未出现缺氧;冬季,DO浓度的垂直分布变化不明显. 阳宗海在秋、夏季形成氧跃层,深水区达到缺氧状态(图3e、h);冬季,DO浓度的垂直分布变化不明显(图3f);春季,形成氧跃层雏形,但深水区暂未出现缺氧现象(图3g). 程海和阳宗海SpCond垂直分布的季节变化如图4a~h所示:程海的SpCond明显高于阳宗海,冬季两湖SpCond的垂直分布相对均一,春季两湖SpCond随水深增加有微弱的上升趋势;秋、夏季,两湖SpCond的垂直分布变化与WT剖面基本呈反向趋势,但夏季程海的SpCond在温跃层升高后随着水深增加而降低直至稳定.

图3 程海(a~d)和阳宗海(e~h)溶解氧浓度垂直剖面的季节变化Fig.3 Seasonality in vertical profile of dissolved oxygen in Lake Chenghai (a-d) and Lake Yangzonghai (e-h)

图4 程海(a~d)和阳宗海(e~h)电导率垂直剖面的季节变化Fig.4 Seasonality in vertical profile of SpCond in Lake Chenghai (a-d) and Lake Yangzonghai (e-h)
2.4 两湖表层水体叶绿素a和营养盐浓度的季节变化
程海秋、冬、春、夏季的表层水体Chl.a浓度分别为5.62±0.92、19.22±11.08、8.06±0.74和12.52±4.70 μg/L(图5a),冬季显著大于秋季(P<0.01)和春季(P<0.05),夏季显著大于春季和秋季(P<0.01),冬季与夏季无显著差异(P>0.05);阳宗海秋、冬、春、夏季的表层水体Chl.a浓度分别为17.64±0.78、45.82±9.41、15.19±0.95和18.27±1.32 μg/L(图5b),冬季显著大于其他3个季节(P<0.01),夏季显著大于春季(P<0.01),夏季与秋季无显著性差异(P>0.05).程海秋、冬、春、夏季的表层水体Chl.a浓度的中位数分别为 5.28、14.39、8.07 和12.57 μg/L,阳宗海的分别为17.55、43.35、15.14 和18.26 μg/L,两湖冬季的值均最高(图5);两湖冬季表层水体Chl.a浓度的变化范围较其他季节更大(图5).
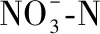
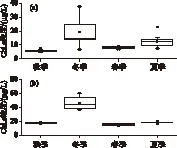
图5 程海(a)和阳宗海(b)表层Chl.a浓度的季节变化 Fig.5 Seasonality in chlorophyll-a concentrations of the surface water column in Lake Chenghai (a) and Lake Yangzonghai (b)

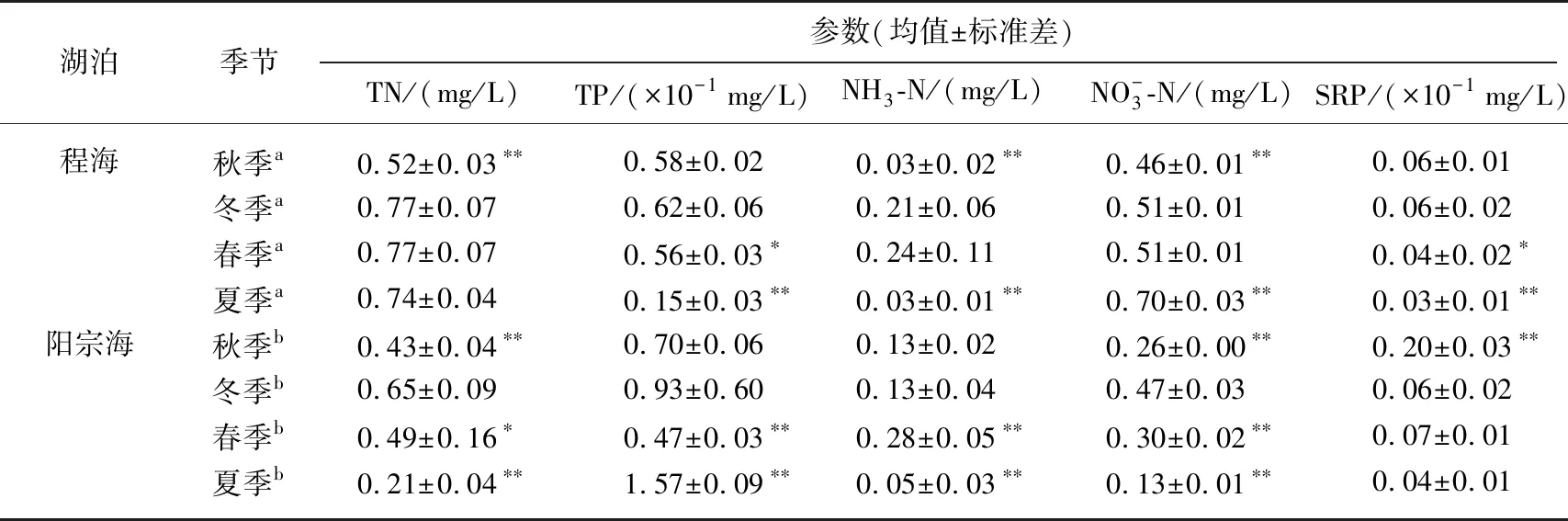
表2 程海和阳宗海表层营养盐浓度的季节变化
aN=9,bN=6;以冬季为参照,*P<0.05,**P<0.01.

表3 四季及季节转化期程海和阳宗海表层Chl.a浓度与有关参数的Spearman相关性
*P<0.05,**P<0.01.
3 讨论
在大尺度空间或长序列时间视野下,湖泊热力分层常受地理位置、湖泊形态、风区长度、辐射、水色及气候变化等因子的影响. 如,虽然高山和亚高山湖泊的平均混合层深度可能相近,但二者的热力结构仍然有差异[33];针对全球26个湖泊(1970-2010年)的研究表明热力分层对气候变化的响应与湖泊形态和平均温度有关[34];针对Lake Pääjärvi和Lake Valkea-Mustajärvi的模拟结果表明,大湖的热力分层更易受风速影响,而小湖更易受辐射或漫射衰减系数的影响[35];Woolway和Merchant认为在未来气候变化背景下诸多湖泊的混合作用会频繁降低[14]. 就温跃层深度而言,本文基于文献[35]中31个湖泊数据的进一步分析表明温跃层深度与湖泊最大水深具有较好的正向线性关系(R2=0.559);针对北温带6个小湖泊的研究表明,水色是影响湖泊热力分层的重要因子,清澈型湖泊的湖上层较深[36];基于千岛湖历史监测数据的研究表明,气候变暖(分层稳定期表层水温升高)和透明度降低使温跃层变浅变厚[10]. 横向比较方面,程海和阳宗海的温跃层深度均较本区域海拔和气象条件相近的抚仙湖浅[16],或与两湖的最大风区长度(或湖区面积)均小于抚仙湖[37]、水深和真光层深度均浅于抚仙湖(最大水深为155.0 m,同期四季真光层深度为14.85±4.54 m)[38]有关;类似地,虽然程海与阳宗海的表层水温、真光层深度并无显著性差异,但程海温跃层的总体位置较阳宗海更深,或与程海的湖区面积和水深均大于阳宗海有关. 风场对湖泊分层尤其是混合层深度具有重要影响[39],不同湖泊的混合层深度随风力的增强而增加. 本研究中,虽然程海和阳宗海的热力分层均存在一定的季节变化,但其四季的混合层深度(分层期以温跃层深度界定,程海最小值为10.25 m,阳宗海最小值为7.33 m)普遍较深,与两湖的有关文献结果接近[21-22],而针对抚仙湖和泸沽湖的研究表明其分层期温跃层深度的最小值大于10 m[16,18];这4个云南湖泊的最小值均大于长江中下游的千岛湖和天目湖[9,10,13],而云南高原的风力强度总体大于长江中下游平原可能是其原因之一. 以阳宗海和千岛湖为例,根据国家气象信息中心资料,1981-2010年云南宜良站(代表阳宗海)1月、4月、7月和10月的累年月平均风速分别为2.0、3.1、1.8 和1.5 m/s,累年月日最大风速≥5.0 m/s日数分别为25.6、28.5、21.1和17.5 d;浙江淳安站(台站编号:58543;代表千岛湖)1月、4月、7月和10月的累年月平均风速分别为1.7、1.6、1.7和1.7 m/s,累年月日最大风速≥5.0 m/s日数分别为7.2、8.9、12.8和5.4 d.
同一湖库热力分层的季节性变化常与辐射、气温、水温、透明度(真光层深度)等涉及水体热量收支的因素有关. 如,太阳辐射被认为是天目湖热力分层变化的主控因子[13],太阳辐射和气温变化对泸沽湖热力分层季节变化具有重要影响[18];针对千岛湖的研究表明透明度和表层水温是千岛湖温跃层深度变化的重要影响因子,其温跃层深度与透明度呈显著正相关、与表层水温呈显著负相关[9]. 程海和阳宗海的表层水温与气温的季节变化趋势一致,与总辐射的变化趋势(两湖所在区域表现出春季最高、夏季次之、冬季最低的趋势)[40]并不完全一致,而两湖热力分层的季节变化表现出了总体一致性与微弱异质性:秋季至冬季,随着太阳辐射减弱、气温降低,水体释放热量,两湖温跃层均消退;冬季至春季,随着太阳辐射与气温升高,表层水温升高,表底层温差增加,两湖均开始出现分层现象;春季至夏季,随着气温升至最高,表底层温差达到最大,使温跃层上移、变厚(程海的温跃层强度变强);夏季至秋季,随着太阳辐射和气温降低,阳宗海的温跃层下移、变薄,但程海的温跃层深度、厚度和强度均未表现出明显变化(可能是因为程海夏秋季的表层水温接近). 由此可见,程海和阳宗海的热力分层均呈现出冬季混合、春季形成分层、夏秋季分层稳定的特征,且未观测到明显的逆温层,说明两湖均属暖单次混合型湖泊[37]. 透明度与真光层深度均可表征水体透光性,透明度(真光层深度)低,表层水体获取的太阳能多、水温高,表层与深层水的温差大,因而透明度降低会使温跃层深度变浅厚度变大[10]、清澈型湖泊的湖上层较浑浊型湖泊更深[36]. 本研究中,两湖的分析结果暗示其真光层深度的单一变化总体上不足以对温跃层深度等参数产生显著影响;当然,这可能与本研究所获取的数据量较少有关,故而并不排除真光层深度或透明度及其与其他因子的联合作用对水体热力分层的影响.
DO是反映水体新陈代谢状况的重要指标,其浓度常受制于大气溶解与(浮游)植物光合作用放氧、水生生物呼吸与有机物降解耗氧等过程的平衡[37]. 热力分层期间,温跃层阻碍了湖上层DO向下扩散,加之湖下层的呼吸分解作用,使湖下层DO浓度减小至缺氧甚至厌氧状态,这在程海和阳宗海热力分层期,尤其在稳定分层的夏秋季表现尤为明显. SpCond可间接表征水体中溶解盐的含量,其值越高表明溶解盐含量越高. 稳定热力分层期(夏秋季),程海和阳宗海的SpCond在温跃层附近均呈现出沿水深升高的趋势,这与淡水贫营养的泸沽湖[18]不同,而与贫中营养的龙滩水库[41]、中营养的天山天池[11]一致;换言之,作为中营养湖泊,程海和阳宗海夏秋季湖下层的溶解盐含量总体较湖上层高,可能与湖下层较强的分解(矿化)作用有关,而关于夏季程海温跃层的SpCond随水深先升高后下降的现象则有待进一步研究. 当秋季转为冬季时,两湖真光层的SpCond均明显升高、底层的SpCond则明显下降,说明随着热力分层的消退,底层的溶解盐可随水体上下层的混合而向表层输送. 加之期间的降水量较少且呈减少趋势,意味着大气沉降与河流输入的营养盐有限,暗示热力分层消退可能是导致表层无机营养盐(如氨氮及富氧条件下经硝化作用产生的硝酸盐氮,亦可能有磷)升高的重要原因(表2). 当冬季转春季时,降水量微弱的升高或不能带来流域营养物质的大量入湖,而可能对水体溶解盐起到一定的稀释作用,进而成为两湖表层及水柱SpCond呈降低趋势的原因之一;从春季的剖面来看,SpCond随水深的增加有微弱的上升趋势,意味着热力分层的初步形成已利于底层溶解盐含量的升高.
需指出的是,本研究主要是基于程海和阳宗海水体分层和浮游植物生物量季节变化特征的初步分析与推断,而缺乏针对周年热力分层转化及其关键时期的水动力、剖面营养盐及浮游植物等数据的长期高频观测,故只是在初步认识两湖水体分层特征的基础上,提供了一种针对浮游植物生物量季节变化,尤其冬季水华发生的案例和可能解释. 而在未来复杂的气候变化与人为干扰背景下,为了更加全面深入地认识程海和阳宗海及本区域其他类似湖泊的水体热力分层特征及其生态环境效应(如,揭示其对浮游植物群落结构变化乃至水华发生的驱动机制等),仍有大量的系统工作需要且值得开展.
4 结论
程海和阳宗海水体分层的特征均为冬季混合、春季形成分层、夏秋季分层稳定,均属暖单次混合型湖泊,程海温跃层的总体位置较阳宗海更深. 热力分层稳定期,两湖水柱底层形成缺氧甚至厌氧条件,底层电导率总体较表层高;秋季转冬季热力分层消退可能是导致水体表层无机营养盐升高的重要原因,加之适宜的光热条件可诱导两湖冬季表层水华的发生(不排除浮游植物垂向迁移的可能贡献);冬季转春季总辐射升至年度最高可能对浮游植物产生光抑制作用,加之两湖电导率(溶解盐浓度)降低及(阳宗海)TP浓度的降低或NH3-N浓度的升高(或有毒性作用),进而使浮游植物生物量降低;春季转夏季,两湖浮游植物生物量的升高不仅与水温升高有关,可能还与辐射降低(光抑制作用减弱)及水柱表层营养盐组分的变化有关. 关于云南乃至云贵高原深水湖泊热力分层特征及其生态环境效应,尤其是对浮游植物群落变化及水华发生影响等的研究,仍有大量的系统性甚至基础性的工作需要开展.
致谢:感谢陈毅良、孔德平、聂菊芬等协助的现场调查或样品分析工作,感谢张运林研究员的帮助!
猜你喜欢
建材发展导向(2022年6期)2022-04-18
煤气与热力(2021年10期)2021-12-02
海洋石油(2021年3期)2021-11-05
建材发展导向(2021年9期)2021-07-16
煤气与热力(2021年4期)2021-06-09
河北环境工程学院学报(2021年1期)2021-03-19
现代装饰(2020年10期)2020-10-28
小学阅读指南·低年级版(2017年2期)2017-03-23
青年文摘·上半月(1989年7期)1989-01-01
