
黄淮北片冬麦区抗赤霉病基因Fhb1种质挖掘及溯源
2020-05-07 08:18张颖君高慧敏李子千胡梦芸孙丽静吕亮杰
华北农学报 2020年2期
张颖君,高慧敏,李子千,胡梦芸,孙丽静,刘 茜,吕亮杰,李 辉
(1.河北省农林科学院 粮油作物研究所,河北省作物遗传育种实验室,河北 石家庄 050035; 2.河北省农林科学院 经济作物研究所,河北 石家庄 050051)
小麦赤霉病(Fusariumhead blight,FHB)是由禾谷镰刀菌引起的一种危害性很强的小麦真菌病害[1]。禾谷镰刀菌在侵染小麦期间,特异诱导次生代谢基因簇fg3_54表达,合成一个非核糖体八肽-镰孢菌素A(Fusaoctaxin A),使菌丝可以从小麦组织内一个细胞穿壁入侵到相邻的细胞,从而导致小麦感病[2]。由于禾谷镰刀菌会产生真菌毒素如脱氧雪腐镰刀菌烯醇(Deoxynivalenol,DON),所以小麦赤霉病不仅影响小麦产量还严重影响小麦加工品质和食用安全性[3-4]。小麦赤霉病在我国长江中下游麦区发生比较严重,如江苏省年均发生面积约120万hm2,超过该省小麦种植面积的50%[5]。而近年来,受气候变化、小麦-玉米轮作和秸秆还田等因素的影响,赤霉病在黄淮冬麦区有加重的趋势[6]。河南省2012年小麦赤霉病发生面积为339.7万hm2,2016年发生面积174.0万hm2 [7]。在河北省中南部小麦赤霉病也有片状发生。因此,提高赤霉病抗性已成为黄淮麦区特别是黄淮南片的主要育种目标之一[6]。黄淮麦区不具备赤霉病田间表型鉴定的理想环境,而人工鉴定多采用单花滴注或田间喷雾接种的方法,费时费力。利用分子标记辅助选择(Marker-assisted selection,MAS)手段进行小麦抗赤霉病种质筛选是培育抗病品种的有效途径。
目前,已定位到的小麦抗赤霉病QTL位点超过100个,以位于小麦3BS染色体的Fhb1基因抗性最强[1,8]。Fhb1基因最初发现于苏麦3和其衍生品种宁7840[9-10],表现为抗病原菌扩展,可降低赤霉病严重度20%左右[11]。多年来人们一直在寻求开发可用于Fhb1基因鉴定的分子标记。Anderson等[12]将Fhb1基因定位到小麦3BS染色体Xgwm533~Xgwm493区间,Cuthbert等[13]和Liu等[14]将其精细定位到标记sts3B-189~sts3B-206之间,遗传距离1.2 cM。Liu等[15]将Fhb1基因缩小到261 kb的区间,开发了共显性标记UMN10。Bernardo等[16]分析了Fhb1基因附近的EST(Expressed sequence tags)序列,在标记Xgwm533~Xgwm493区间发现7个SNP与小麦赤霉病抗性相关,其中Xsnp3BS-8存在于苏麦3号及其衍生品种中。Rasheed等[17]根据Xsnp3BS-8位点开发了KASP(Kompetitive allele specific PCR)标记用于小麦种质材料Fhb1基因检测。Rawat等[18]克隆了PFT(Pore-forming toxin-like)基因,认为PFT可能是Fhb1基因,PFT有3种基因型:PFT-Ⅰ(苏麦3号类型)、PFT-Ⅱ(南大2419类型)和PFT-Ⅲ(基因缺失类型)[6]。这些标记在小麦抗赤霉病育种过程中起到了一定的作用,但不能将Fhb1等位基因有效区分。Su等[19]和朱展望等[6]分析了PFT邻近的His(Histidine-rich calcium-binding protein)基因序列,发现在苏麦3号类型品种中His基因在5′-UTR区(Untranslated region)和部分ORF区(Open reading frame)存在序列缺失。His基因有3种等位基因类型,分别为His-Ⅰ(苏麦3号类型,1 309 bp)、His-Ⅱ(2 061 bp)和His-Ⅲ(2 061 bp)。其中His-Ⅰ起始密码子附近有752 bp缺失,His-Ⅲ与His-Ⅱ长度相同但存在4个SNP变异。根据此InDel区段Su等[19]开发了共显性标记TaHRC-GSM和KASP标记TaHRC-Kasp,朱展望等[6]开发了共显性标记His-InDel,这些标记可以较有效的区分小麦赤霉病抗/感种质。本研究对336份小麦主栽品种和种质材料利用KASP标记TaHRC-Kasp进行检测,然后利用His基因的共显性标记His-InDel进行进一步验证,以期筛选出含有Fhb1基因的种质材料,为黄淮冬麦区抗赤霉病小麦育种提供宝贵的遗传信息及亲本材料。
1 材料和方法
1.1 供试材料
本研究共选择336份小麦主栽品种和种质作为试验材料,其中黄淮冬麦区312份(包括河北136份、河南89份、山东72份、陕西15份)。由于江苏、安徽两省是小麦赤霉病的重发区,所以选择了24份材料(江苏17份、安徽7份)一并进行Fhb1基因检测。
1.2 KASP标记检测
根据Su等[19]开发的Fhb1基因的KASP标记,在抗赤霉病基因型(Fhb1+)引物添加FAM荧光,在感赤霉病基因型(Fhb1-)引物添加HEX荧光。Fhb1+引物序列为5′-GAAGGTGACCAAGTTCATGCTTTGGGCTCACGTCGTGCAAATGGT-3′,Fhb1-引物序列为5′-GAAGGTCGGAGTCAACGGATTTGTCTGTTTCGCTGGGATG-3′,反向通用引物序列为5′-CTTCCAGTTTCTGCTGCCAT-3′。利用此KASP标记对336份小麦材料进行检测,采用384孔PCR板,3 μL反应体系(包括DNA模板60~100 ng,2×KASP master mixture 2 μL,引物混合液0.056 μL),PCR扩增程序为94 ℃预变性15 min;94 ℃变性20 s,65~57 ℃(每循环降温1 ℃)退火1 min,10个循环;94 ℃变性20 s,57 ℃退火1 min,35个循环。KASP检测采用LGC公司(http://www.lgcgenomics.com)SNP分型检测系统完成。
1.3 His-InDel标记验证及His基因序列分析
采用朱展望等[6]开发的His-InDel标记对KASP检测的部分材料进行验证,His-InDel标记正向引物序列为5′-ATGCGTGCGCTGTACTTG-3′,反向引物序列为5′-CGTCACAGAGTCCAGTGAAA-3′。PCR反应体系:2×PCR Mix 10 μL,DNA模板约100 ng,上下游引物(10 μmol/L)各1 μL,加ddH2O至20 μL。PCR Mix购于康为世纪有限公司(http://www.cwbiotech.com/),引物由英潍捷基(上海)贸易有限公司(http://invitrogen.tecenet.com)合成。PCR扩增在BioRad公司(http://www.bio-rad.com)MyCycler PCR仪上进行,扩增程序为95 ℃预变性3 min;94 ℃变性30 s,65 ℃退火30 s,72 ℃延伸2 min 30 s,共35个循环;最后72 ℃ 延伸7 min,10 ℃保温。PCR产物用1%琼脂糖凝胶电泳进行检测,溴化乙啶(Ethidium bromide,EB)浸泡染色后在BioRad公司凝胶成像系统下照相,记录电泳结果。目标条带用琼脂糖凝胶回收试剂盒(天根生化科技有限公司,http://www.tiangen.com)进行回收,操作步骤参见试剂盒说明书。T-载体采用北京全式金生物技术有限公司(http://www.transgen.com.cn)Easy-T3载体,连接体系为T-载体1 μL,回收产物4.0 μL,室温连接20 min。连接后的载体利用热激法转化大肠杆菌感受态细胞DH5α(北京全式金生物技术有限公司),转化后的大肠杆菌DH5α涂布在含氨苄青霉素(100 mg/L)的LB培养基(酵母提取物5 g、胰蛋白冻10 g、NaCl 10 g、琼脂粉6 g、加蒸馏水至1 L)上,并进行蓝白斑筛选。每个LB平板挑取6个阳性克隆进行测序,序列测定由深圳华大科技公司(http://www.bgitechsolutions.cn)完成,序列分析采用软件DNAMAN(http://www.lynnon.com)和Vector NTI Advance 10(http://www.invitrogen.com)进行。
1.4 PCA分析
对供试的336份材料采用小麦15K芯片(中玉金标记生物技术股份有限公司,http://www.cgmb.com.cn)进行SNP位点检测。利用R语言编写程序对SNP数据进行质控(Quality control),去除数据缺失(Missing data)>0.5、杂合率>0.1和最小基因频率(Minmum allele frequency,MAF)>0.05的数据。利用Tassel 5.0 软件进行PCA(Principal component analysis)分析。
2 结果与分析
2.1 Fhb1基因KASP标记检测结果
利用Fhb1基因KASP标记对供试的336份小麦材料进行检测,有2个河北材料(石家庄75、紫茎白)与3个来自江苏的抗赤霉病材料(苏麦3号、宁麦9号和宁麦13)携带FAM荧光,聚集在y轴附近,表现为抗赤霉病类型(Fhb1+);有3个材料(宁麦16、德宏福6号和泊麦7号)表现为杂合基因型(Fhb1+/-);其余材料携带HEX荧光信号,聚集在x轴附近,为感赤霉病类型(Fhb1-)(图1)。
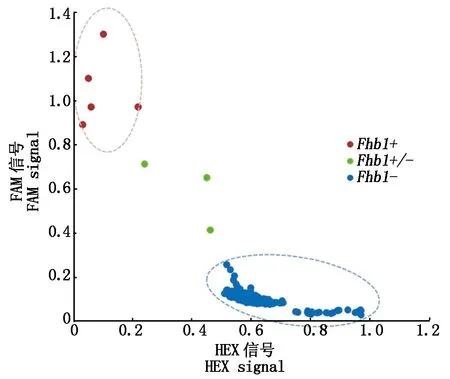
图1 Fhb1基因KASP标记检测结果Fig.1 Fhb1 genotyping analysis using KASP Marker
2.2 His-InDel标记检测结果
为了验证KASP标记的准确性,采用朱展望等[6]开发的His-InDel标记对其进行进一步检测。检测品种包括5个KASP标记检测显示抗赤霉病类型的材料(石家庄75、紫茎白、苏麦3号、宁麦9号和宁麦13)、3个杂合基因型材料(宁麦16、德宏福6号和泊麦7号)和16个感赤霉病类型材料(郑麦9023、小偃22等)。从图2可以看出,His-InDel为共显性标记,可以将材料进行明显区分。石家庄75、紫茎白、苏麦3号、宁麦9号、宁麦13和宁丰518扩增得到1 309 bp条带,表现为His-Ⅰ基因型(抗赤霉病类型);郑麦9023、西农979、小偃22等17个材料扩增得到2 061 bp条带,为His-Ⅱ(感赤霉病类型);宁麦16表现为Fhb1杂合类型。KASP标记与His-InDel标记检测结果基本一致,但宁丰518品种KASP检测为Fhb1-基因型,而His-InDel标记检测为Fhb1+基因型;德宏福6号和泊麦7号2份材料KASP检测为杂合类型,而His-InDel标记检测为Fhb1-基因型。
2.3 His基因序列分析结果
对苏麦3号、石家庄75和紫茎白3个品种的His-InDel标记PCR扩增条带进行回收、测序,分析其His基因序列。结果显示,石家庄75、紫茎白与苏麦3号的His基因序列一致,为1 309 bp,表现为Fhb1+基因型(图3)。测序结果进一步验证了His-InDel标记检测的准确性。

1.郑麦9023;2.西农979;3.小偃22;4.郑麦366;5.冀5265;6.冀麦325;7.济麦22;8.矮抗58;9.农大5363;10.轮选103;11.德宏福6号;12.泊麦7号;13.周麦35;14.豫教黑1号;15.晋麦8号;16.宁麦13;17.宁麦16;18.皖981;19.皖农306;20.宁丰518;21.石家庄75;22.紫茎白;23.苏麦3号;24.宁麦9号;M.Marker Ⅲ。
1.Zhengmai 9023;2.Xinong 979;3.Xiaoyan 22;4.Zhengmai 366;5.Ji 5265;6.Jimai 325;7.Jimai 22;8.Aikang 58;9.Nongda 5363;10.Lunxuan 103;11.Dehongfu 6;12.Bomai 7;13.Zhoumai 35;14.Yujiaohei 1;15.Jinmai 8;16.Ningmai 13;17.Ningmai 16;18.Wan 981;19.Wannong 306;20.Ningfeng 518;21.Shijiazhuang 75;22.Zijingbai;23.Sumai 3;24.Ningmai 9;M.Marker Ⅲ.
图2 His-InDel标记PCR检测结果
Fig.2 PCR results of His-InDel Marker

图3 不同品种His基因序列比对分析Fig.3 Alignment of His gene sequences of different cultivars
2.4 基于全基因组SNP位点分析供试小麦材料的亲缘关系
利用小麦15K芯片对336份材料进行检测,共获得SNP位点13 947个。通过数据质控,去除4 168个位点,最终获得9 779个SNP位点用于供试材料的PCA分析。从PCA结果(图4)可以看出,336份小麦材料主要分为3个群,分别为河北(红色菱形)、河南(绿色三角)和山东(蓝色圆圈)。来自陕西(橙色圆圈)的材料多数与来自河南的材料聚在一起,来自江苏(紫色方块)和安徽(黄色菱形)的材料多数聚在坐标轴中心附近。来自河北的携带Fhb1基因的2个材料石家庄75和紫茎白位于坐标轴中心附近,这2个材料与来自江苏的材料亲缘关系较近。

图4 336份小麦材料PCA分析结果Fig.4 PCA analysis of 336 wheat samples
3 结论与讨论
小麦赤霉病田间表型鉴定受天气和接种方法等多种因素影响,稳定性和重复性差,经常需要进行多年多地点鉴定才能提高鉴定的准确性[20],而黄淮麦区因缺少适合田间表型鉴定的气候条件,所以赤霉病抗性品种的筛选难度更大。因此,利用分子标记辅助选择(MAS)可以大大提高赤霉病抗性品种的培育速度。Fhb1是目前为止报道的效应最大的赤霉病抗性基因,前人一直致力于Fhb1基因的克隆和标记开发以服务于小麦抗赤霉病育种[6,12-19]。本研究利用Su等[19]开发的Fhb1基因的KASP标记TaHRC-Kasp对来自黄淮冬麦区(河北、河南、山东、陕西)和江苏、安徽的共336份小麦材料进行检测,共检测出5份携带Fhb1基因的品种,其中有2份是来自河北省的品种石家庄75和紫茎白,其余3份为来自江苏省的苏麦3号、宁麦9号和宁麦13,这3份材料是公认的携带Fhb1基因的品种。有3份材料(宁麦16、德宏福6号和泊麦7号)表现为Fhb1杂合基因型,其余材料为Fhb1-基因型。为了验证KASP标记检测结果,本研究挑选了24份材料利用朱展望等[6]开发的His-InDel标记做进一步检测。His-InDel标记与KASP标记检测结果基本一致,但宁丰518品种KASP检测为Fhb1-基因型,而His-InDel标记检测为Fhb1+基因型;德宏福6号和泊麦7号2份材料KASP检测为杂合类型,而His-InDel标记检测为Fhb1-基因型,这可能是受KASP标记检测的特异性或荧光信号的偏差所影响。KASP标记由于检测速度快、通量高,已广泛应用于作物分子标记辅助育种。从本研究结果看,TaHRC-Kasp标记基因分型清晰,检测准确性也较高,将TaHRC-Kasp标记和His-InDel标记结合使用可以大大提高小麦抗赤霉病育种效率。
Fhb1基因在我国江苏、安徽等赤霉病重发区已得到广泛应用,并且在北美和日本应用也较为广泛[18,20-21]。研究表明,中国小麦品种所含Fhb1基因主要有2个来源,分别为苏麦3号和宁麦9号,利用苏麦3号在长江中下游麦区育成了宁7840等品种,利用宁麦9号培育成了宁麦系列品种(如宁麦13、宁麦14、宁麦26等)和扬麦系列品种(如扬麦18、扬麦21等)[6]。随着近年来黄淮麦区赤霉病呈加重趋势,提高小麦赤霉病抗性已成为黄淮南片麦区主要育种目标之一,但Fhb1基因在黄淮麦区的应用相对缓慢,目前尚无携带Fhb1基因的小麦品种报道。究其原因一则可能与黄淮麦区抗赤霉病育种时间较短有关,二则可能是由于Fhb1基因与某些不利农艺性状连锁较紧密[6],导致在育种进程中Fhb1基因受到人工选择压力而被去除。有研究表明,Fhb1+基因型与秆锈病基因Sr2-基因型紧密连锁[22],在很大程度上限制了Fhb1基因的应用,本研究也发现6个Fhb1+基因型品种(石家庄75、紫茎白、苏麦3号、宁麦9号、宁麦13和宁丰518)不携带Sr2抗性基因。由于光温等气候条件的差异,长江中下游春麦区的抗赤霉病小麦品种很难在黄淮冬麦区直接应用,因此,在黄淮冬麦区筛选携带Fhb1基因的种质材料对黄淮冬麦区抗赤霉病小麦育种尤为重要。本研究利用TaHRC-Kasp和His-InDel标记筛选出2个来自河北省的材料(石家庄75和紫茎白)携带Fhb1基因。His基因序列分析也显示石家庄75、紫茎白和苏麦3号、宁麦9号、宁麦13、宁丰518的序列一致,为抗赤霉病类型;郑麦9023、小偃22、矮抗58、泊麦7号等为感赤霉病类型。小麦品种石家庄75的亲本组合为“矮立多/碧玉麦//安徽5号”,矮立多(Ardito)是从意大利引进的品种,碧玉麦(Quality)是20世纪40年代初从美国引进的春麦材料,矮立多和碧玉麦不具有Fhb1基因。安徽5号的亲本为南大2419和胜利麦。南大2419是从1932年引入的意大利品种Mentana中混选出来的,胜利麦是建国初期的农家品种。南大2419是Fhb1-材料,所以,石家庄75中的Fhb1基因有可能是来源于胜利麦,但这还需进行进一步验证核实。紫茎白是河北省的地方农家种,其携带Fhb1基因,这说明最初Fhb1基因在国内一些地方品种中有所分布(如望水白、白三月黄、黄方柱、火烧麦、火烧白日麦和菜籽黄等)[23-26],但在现代育种进程中受到人工选择的压力而逐渐消失。
经TaHRC-Kasp和His-InDel标记检测和His基因序列分析,证明来自河北省的2份小麦材料石家庄75和紫茎白携带Fhb1基因。TaHRC-Kasp和His-InDel标记联合使用进行分子标记辅助选择可以大大提高抗赤霉病小麦育种效率。
致谢:感谢中国农业科学院作物科学研究所何中虎研究员、夏先春研究员在Fhb1基因KASP标记检测过程中提供的帮助,感谢湖北省农业科学院粮食作物研究所朱展望博士提供His-InDel标记信息。
猜你喜欢
传染病信息(2022年4期)2022-11-23
今日农业(2022年16期)2022-11-09
农业知识(2022年9期)2022-10-13
西部医学(2022年9期)2022-09-26
今日农业(2021年8期)2021-11-28
今日农业(2021年9期)2021-11-26
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
今日农业(2021年9期)2021-07-28
生物学教学(2018年8期)2018-09-03
