
谷子、小麦籽粒蛋白、淀粉构成及结构差异分析
2020-05-07 04:55赵子龙曹婷婷李素敏王从磊时晓伟杨晓然王建贺李素英
华北农学报 2020年2期
刘 丹,赵子龙,梁 丹,曹婷婷,李素敏,王从磊, 时晓伟,杨晓然,张 明,朱 铭,王建贺,冯 刚,李素英
(1.天津市农作物研究所,天津市农作物遗传育种重点实验室,天津 300384;2.唐山师范学院 生命科学系,河北 唐山 063000)
谷子起源于我国,已有8 700年的栽培历史,营养均衡,历史上曾是养育中华民族的主粮。目前,谷子主要分布于中国、印度等发展中国家,中国谷子种植面积占全球的80%左右。小麦起源于亚洲西部,公元前2 000年左右,我国开始种植小麦[1]。由于小麦适口性突出,逐步发展为全球性主粮,在我国也逐步取代谷子的主粮地位;而谷子虽然具有营养全面、均衡,但由于适口性较差,逐步退出主粮地位、成为杂粮,也成为制约其产业发展的主要限制因素。
蛋白质、淀粉是作物籽粒的主要营养成分,二者共同决定了作物的品质及产量,对作物具有重要意义[2-3]。小麦中的蛋白可形成面筋,用于制作面包、面条、馒头等食物[4],而谷子、水稻、玉米中籽粒的成分基本不能形成面筋,难以单独制作面包、面条、馒头等食物。淀粉为多聚葡萄糖,占籽粒成分的80%左右,依据糖苷键的不同分为直链淀粉和支链淀粉两类[5],两类淀粉均以颗粒状形态积累在胚乳中,体积大的淀粉颗粒为A型淀粉粒,体积小的淀粉颗粒为B型淀粉粒,体积最小的淀粉颗粒为C型[6-8]。在扫描电镜下,淀粉粒呈现圆球形、椭球形、多面形,且不同作物A型、B型、C型淀粉颗粒含量明显不同。作物籽粒中的蛋白质占籽粒成分的10%~15%,依据溶解性的不同,分为清蛋白、球蛋白、醇溶蛋白和谷蛋白,其中清蛋白为水溶性蛋白,球蛋白为盐溶性蛋白,醇溶蛋白主要溶解于醇类物质,而谷蛋白主要溶解于酸或碱[9-10]。不同作物间蛋白种类及其构成比例和构成蛋白质的氨基酸种类和含量存在较大差异[11-13]。因此,如何通过研究籽粒蛋白、淀粉构成及结构,改良谷子品质,成为摆在广大农业科研工作者面前的一个重要课题,也是解决制约谷子产业发展的瓶颈难题的关键所在。
关于小麦蛋白成分及其构成比例已有一些报道。Henkrar等[14]研究表明,小麦籽粒中,麦谷蛋白和醇溶蛋白均占蛋白总量的40%左右,二者通过蛋白质之间的相互作用形成面筋。谷蛋白决定了面团的弹性,弹性好的小麦粉,即强筋小麦粉适合制作面包,而弹性小,延展性好的面粉,即弱筋小麦粉,适合制作饼干[15-18]。因此,谷蛋白与小麦的营养品质及加工品质密切相关;麦谷蛋白为复合体蛋白,各个亚基之间通过分子内及分子间二硫键、氢键连接聚合,通过巯基乙醇及二硫苏糖醇等还原剂可形成单体蛋白,这些单体蛋白按照其在SDS-PAGE电泳中的迁移速率可分为高分子量谷蛋白亚基(HMW-GS)和低分子量谷蛋白亚基(LMW-GS)[19-21]。醇溶蛋白(Gliadin)作为面筋的主要成分之一,决定了面筋的黏性和延展性[22],不同于麦谷蛋白,Gliadin为单体蛋白,分子量介于20~100 ku,分子间没有二硫键,依托分子内二硫键、氢键及疏水作用力形成球状结构[22]。根据醇溶蛋白在A-PAGE电泳图谱上的相对迁移速率可以将醇溶蛋白分为α、β、γ、ω4类,其中α型醇溶蛋白占Gliadin总量的25%,β型醇溶蛋白占Gliadin总量的30%,γ型醇溶蛋白占Gliadin总量的30%,ω型醇溶蛋白占Gliadin总量的15%[23-25]。然而,目前,对于谷子籽粒蛋白的研究相对较少,也缺乏研究的深度和广度。
关于小麦淀粉成分及其构成比例也有一些研究。小麦籽粒中80%以上为淀粉,淀粉质体为其合成的场所,在高等植物中存在的原质体、叶绿体、淀粉质体相互转化的现象,同时也伴随着淀粉的合成与降解[26]。有研究认为,淀粉质体是由原质体和叶绿体发育而来,原质体和叶绿体中的淀粉积累充分后,二者便转化为淀粉质体;淀粉质体合成的淀粉粒为A型淀粉粒,A型淀粉粒通过自身生长的小淀粉粒发育成B型淀粉粒,对于C型淀粉粒的来源目前尚无研究证实[8, 11]。A型淀粉粒占籽粒淀粉数量的5%左右,质量为籽粒总淀粉量的70%,B型淀粉粒占籽粒淀粉粒总量的95%,但是质量仅占籽粒淀粉总量的25%~30%[8, 11]。同时表达谱分析表明,A型淀粉粒出现在开花后的4~14 d,B型淀粉粒则出现在开花后14 d到籽粒成熟,说明B型淀粉粒可能来源于已经发育成熟的A型淀粉粒。同时A型淀粉粒和B型淀粉粒对于品质的贡献也不相同,A型淀粉粒含量高的小麦制作的面包体积较大,质量较好。然而,目前,对于谷子籽粒淀粉的研究相对较少,对于品质贡献方面的研究也鲜有报道。
谷子和小麦的淀粉、蛋白质结构、形状以及成分均存在较大差别,这也是造成它们营养品质、加工品质以及适口性等差别的主要原因,也是造成谷子逐步退出主粮地位以及制约其产业发展的关键所在。本研究利用扫描电镜对小麦、谷子籽粒结构的进行观察,探明小麦和谷子中淀粉、蛋白质的形态差异及排布特征,并通过对谷蛋白和醇溶蛋白的提取,研究谷蛋白和醇溶蛋白在分子量及组成上的差异,为探明谷子和小麦品质间差异形成的原因提供理论依据,为进一步通过分子手段改良小米品质提供技术支撑,为拓宽谷子品质改良的途径提供新的思路。
1 材料和方法
1.1 试验材料
供试谷子材料由唐山师范学院生命科学系提供,供试小麦材料由天津市农作物研究所小麦中心提供(表1)。

表1 材料Tab.1 Materials
1.2 谷蛋白提取
试验在唐山师范学院进行,采用同时提取高分子量谷蛋白和低分子量谷蛋白的方法对小麦的谷蛋白进行提取,取1粒小麦种子磨碎,将粉末置于2.0 mL离心管中,加入800 μL Buffer A (7.5%异丙醇; 0.3 mol/L NaI),于65 ℃提取1 h,其间10 min用振荡仪振荡1次后10 000 r/min,离心3 min,弃上清,并重复1次;在沉淀中加入200 μL Buffer B(25% 异丙醇,0.04 mol/L Tris-HCl(pH=8.0),10%SDS,2%DTT),涡旋充分后,于65 ℃提取1 h,其间不断摇动(10 min涡旋1次);再加入200 μL Buffer C (25%异丙醇,0.04 mol/L Tris-HCl(pH=8.0),10%SDS,1.4%乙烯基吡啶),涡旋混匀,于65 ℃提取30 min,10 000 r/min,离心5 min;将上述400 μL的上清吸取出来,转移到另一干净离心管中,加入200 μL Buffer D(0.625 mol/L Tris-HCl(pH=6.8),5%硫基乙醇,20%甘油,2%SDS,0.2%溴酚蓝),涡旋混匀,静置30 min后,放入沸水中100 ℃,变性5 min,自然冷却,该溶液即为含有HMW-GS和LMW-GS的提取液,可做电泳分析用。通过SDS-PAGE,浓缩胶浓度为10%,分离胶浓度为5%,稳流15 mA/板,电泳16 h,对小麦谷蛋白进行鉴定。
通过植物总提蛋白方法,对谷子冀谷32进行籽粒蛋白总提(提取Buffer:0.625 mol/L Tris-HCl,5%硫基乙醇,20%甘油,2% SDS,0.2%溴酚蓝;涡旋混匀,静置30 min),通过60%异丙醇去除醇溶蛋白的方法对谷蛋白进行提取,并通过SDS-PAGE对谷子总蛋白、谷蛋白进行鉴定。
1.3 醇溶蛋白提取
采用70%乙醇对春小麦、冬小麦醇溶蛋白进行提取,采用ISTA 1986年颁布的A-PAGE电泳流程对小麦醇溶蛋白进行鉴定[27];采用65%异丙醇对谷子籽粒进行醇溶蛋白提取,并通过A-PAGE对谷子醇溶蛋白进行鉴定。
1.4 籽粒电镜观察
选择外形完整、颗粒饱满的谷子、小麦籽粒,各种籽粒随机选取3粒。将干燥好的籽粒,用徒手切片法切下断裂面,厚度约为1~2 mm,即为样品。将样品自然断裂面朝上,切割面朝下,用导电胶粘着于样品台上,然后将样品台置于样品室放在场发射扫描电镜(ZEISS Sigma 300)下,依次观察各个籽粒蛋白断裂面的腹部、中部、背部,并拍照记录。
2 结果与分析
2.1 谷子与小麦谷蛋白的对比分析
2.1.1 小麦谷蛋白SDS-PAGE分析 SDS-PAGE电泳分析结果表明:小麦品种中国春、津强8号、津强9号、津强11号、津强12号、106、CB037、轮选987、津硬8号中均存在高分子量谷蛋白亚基和低分子量谷蛋白亚基,但高分子量谷蛋白亚基条带较少,一般表达4~5种高分子量谷蛋白亚基,而低分子量谷蛋白亚基组成较为复杂,品种间变异较为丰富(图1)。前人研究表明,中国春的高分子量组蛋白为Dx2+Dy12和Bx7+By8,A基因组上的高分子量谷蛋白呈沉默状态,不表达[28]。津强8号高分子量谷蛋白表达模式与中国春类似,仅4种高分子量谷蛋白表达,而津强9号、津强11号、津强12号、106与中国春相比,除了B基因组和D基因组的高分子量谷蛋白表达,A基因组上也有一个位点表达产生了高分子量谷蛋白,因此这些品种具有5条高分子量谷蛋白条带(图1)。CB037和轮选987B组染色体上谷蛋白与中国春存在明显差异,Bx和By的分子量接近。津硬8号为四倍体小麦,仅含有A基因组和B基因组,但其高分子量谷蛋白也有4个位点表达(图1),说明在进化过程中,四倍体小麦的A基因组上的高分子量谷蛋白编码基因并未沉默。低分子量谷蛋白编码位点较多,且表达具有品种间特异性,但目前对于低分子量谷蛋白分布及功能的研究较少。
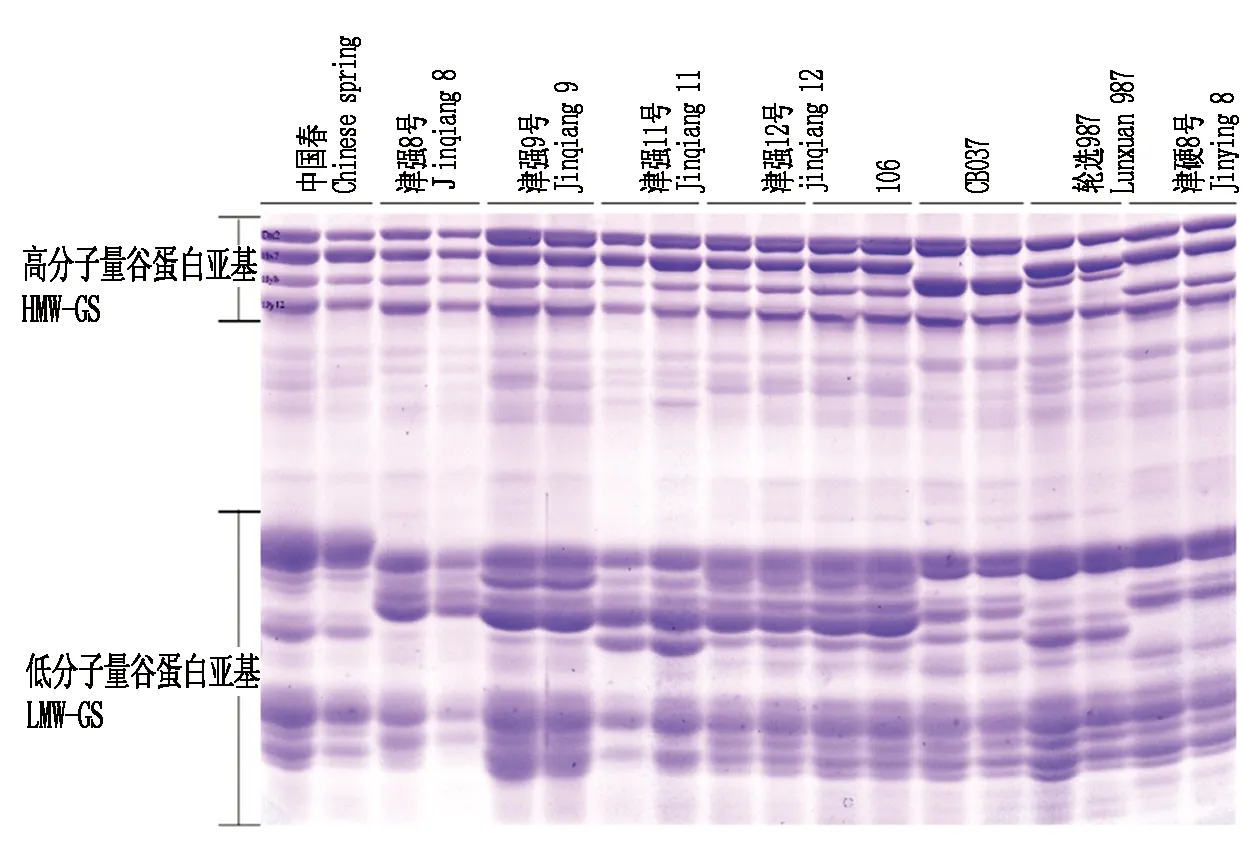
图1 小麦谷蛋白亚基Fig.1 Wheat gluten subunit
2.1.2 谷子谷蛋白SDS-PAGE分析 对谷子总蛋白和谷蛋白的分析结果(图2)表明:谷子新品种—冀谷320.12,0.15,0.18,0.21,0.24 g籽粒提取的蛋白浓度逐渐上升,0.12 g谷子籽粒所提取的蛋白与0.015 g小麦籽粒提取的总蛋白亮度接近,说明谷子籽粒蛋白质含量不如小麦含量丰富,同时对谷子谷蛋白提取发现,谷子谷蛋白含量很低,仅有1条明显的谷蛋白条带和2条较弱的谷蛋白条带。
对比小麦总蛋白带谱发现,谷子总蛋白的迁移速率显著低于小麦总蛋白迁移速率,说明谷子蛋白的分子量显著低于小麦蛋白分子量。

1.小麦(中国春),0.015 g籽粒总蛋白;2-6. 0.12,0.15,0.18,0.21,0.24 g谷子籽粒总蛋白;7-11. 0.12,0.15,0.18,0.21,0.24 g谷子籽粒谷蛋白。
1.Wheat(Chinese spring), 0.015 g grain for total protein extraction; 2-6. 0.12, 0.15, 0.18, 0.21, 0.24 g grain for total protein extraction of foxtail millet; 7-11. 0.12, 0.15, 0.18, 0.21, 0.24 g grain for glutenin extraction of foxtail millet.
图2 谷子总蛋白及谷蛋白
Fig.2 Protein of foxtail millet
2.2 谷子与小麦醇溶蛋白的对比分析
2.2.1 小麦醇溶蛋白分析 通过单粒小麦种子(约0.06 g),进行醇溶蛋白提取,并利用酸性聚丙烯酰胺凝胶电泳(A-PAGE)对醇溶蛋白进行分析,发现在小麦的醇溶蛋白在ω、γ、β、α区均有分布,且醇溶蛋白含量丰富,组成复杂,每个品种均具有自身的条带构成特征,且变异丰富(图3)。中国春与Fielder在ω区存在差异,Fielder ω区有4个位点表达,中国春有5个位点表达;γ区Fielder表达的位点多于中国春;α/β区,2个品种的表达相似。济南17与Fielder在ω区醇溶蛋白组成相似,但是在α/β区存在显著差异;济南44与中国春在ω区醇溶蛋白组成相似。总体说明,醇溶蛋白的组成具有品种间特异性(图3-A)。对于天津市农作物研究所的冬麦资源分析发现,冬麦材料的醇溶蛋白在ω、γ、β、α区均存在明显差异,品种特异性差异显著(图3-B)。说明,小麦中醇溶蛋白具有明显的品种间特异性。
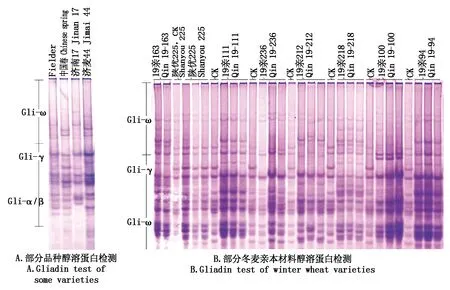
图3 小麦醇溶蛋白Fig.3 Gliadin of wheat varieties
2.2.2 谷子醇溶蛋白分析 通过提取谷子醇溶蛋白(约0.06 g),进行A-PAGE电泳分析(图4)表明,铁谷8(V1)、兴谷88(V2)、坝谷214(V3)、朝谷13(V4)、延谷12(V5)、公谷70(V6)、赤谷4(V7)、大同29(V8)、郑9188(V9)品种中,均含一定量的醇溶蛋白,谷子醇溶蛋白约10~20条条带,在不同品种间存在多态性,条带种类和组成具有品种特征。V1中含有12个醇溶蛋白条带,其中有2个条带表达量较高,其余条带表达较弱;V2有9条醇溶蛋白条带,表达量较高的条带与V1品种的条带位置不同;V3、V4、V9的醇溶蛋白带谱相似;V5、V7醇溶蛋白带谱相似,醇溶蛋白条带亮度低,说明表达丰度较低;V8有3条高亮度条带。说明谷子不同品种醇溶蛋白含量可能存在一定差异,并且醇溶蛋白存在品种间特异性。

图4 谷子醇溶蛋白Fig.4 Alcohol-soluble protein of foxtail millet
2.3 籽粒电镜观察
通过扫描电镜观察表明,谷子和小麦籽粒中均含有淀粉粒和蛋白体。在常规谷子品种冀谷32、恢复系JK4-227、不育系51A中,均含有两类淀粉粒,一类体积较大,成多边形,直径5~10 μm,属于B型淀粉粒,含量丰富;另一类体积较小,为球形,直径约2 μm,属于C型淀粉粒,含量少于B型淀粉粒;在淀粉粒间隙填充着较少的蛋白体。在春小麦津强1号、冬小麦津农10号、轮选987中存在三类淀粉粒,大小不同,其中,一类淀粉粒体积较大,直径10~30 μm,呈盘子状,为A型淀粉粒,含量丰富;第二类淀粉粒体积比A型淀粉粒小,直径5~10 μm,成球形或者多边形,为B型淀粉粒;第三类淀粉粒体积最小,直径小于2 μm,为C型淀粉粒,B型、C型淀粉粒显著少于A型淀粉粒;同时淀粉粒间隙中填充着较为丰富的蛋白体,不同品种淀粉体覆盖的蛋白含量不同,津农10号淀粉体表面较为平滑,而津强1号淀粉粒表面被蛋白体紧密包裹(图5)。

谷子冀谷32为常规品种、JK4-227为恢复系、51A为不育系;小麦津强1号(春小麦,强筋)、 津农10号(冬小麦,中筋)、轮选987(冬小麦,中筋)。 Jigu 32 was a conventional variety of foxtail millet, JK4-227 was a restorer line, and 51A was a sterile line; Jinqiang 1 (spring wheat, strong-gluten), Jinnong 10 (winter wheat, middle-gluten), Lunxuan 987 (winter wheat, middle-gluten).
3 讨论
小麦面筋含量高,因此可用于制作面包、面条、蛋糕等食品;而谷子几乎不含有面筋,但营养丰富。因此,可以以小米面主要作为配粉,按照一定比例,添加到小麦粉中,制作面包、面条、蛋糕,既可增加这些产品的营养价值,又可提高小米的适口性和商业价值,从而扩大小米的用量,解决制约谷子产业发展的小米用途窄的难题。
据报道,小麦籽粒蛋白含量约13%[29],而谷子的蛋白含量为10%[30]。本研究发现,谷子不仅总蛋白含量低于小麦,在SDS-PAGE上,谷子总蛋白迁移速率显著快于小麦蛋白迁移速度,说明谷子籽粒蛋白的分子量显著低于小麦总蛋白。小麦与谷子的蛋白质含量差异不大,不同品种的小麦籽粒中均含有谷蛋白和醇溶蛋白,二者共同形成面筋。本研究也证实,小麦品种都含有高分子量谷蛋白亚基和低分子量谷蛋白亚基,而谷子的籽粒中谷蛋白很少,且与小麦相比,谷蛋白的分子量很低,说明谷子含有谷蛋白的水平很低,并且都是一些分子较小的谷蛋白,与小麦谷蛋白存在明显差异。醇溶蛋白研究发现,谷子的醇溶蛋白条带较少,多态性不如小麦籽粒醇溶蛋白丰富,说明谷子中编码醇溶蛋白的基因少于小麦,在改良谷子品质时,可以适当引入分子量大的谷蛋白、醇溶蛋白亚基,增加谷子蛋白的分子量。
谷子中的谷蛋白和醇溶蛋白含量与小麦存在较大差异,说明二者蛋白质性质和构成差异均较大,谷子的蛋白质含量虽然接近小麦,但是由于其谷蛋白和醇溶蛋白含量少,且分子量与小麦存在较大差异,所以不能形成面团,制作面包等商品。
研究发现,小麦籽粒蛋白中20%为清蛋白和球蛋白,其余80%为谷蛋白和醇溶蛋白,谷蛋白和醇溶蛋白的比例接近1∶1[10, 31]。本研究发现,从带谱图上看,谷子含有一定的醇溶蛋白,也具有一定的多态性,但是谷子谷蛋白含量极低,分子量也较小,这说明谷子醇溶蛋白的含量远远高于谷蛋白含量,与小麦谷蛋白和醇溶蛋白的比例差异较大,也暗示由于谷子谷蛋白和醇溶蛋白含量差异悬殊,因此不能形成面筋。
谷子、小麦除了蛋白质存在差异外,在淀粉粒上也存在较大差异。小麦主要存在A、B、C三类淀粉体,其中A型淀粉粒体积较大,直径10~30 μm,呈盘子状;B型淀粉粒体积比A型淀粉粒小,直径5~10 μm,呈球形或者多边形;C型淀粉粒体积最小,直径小于10 μm,与蛋白体镶嵌在一起[32]。谷子的淀粉体大部分呈多边形,直径约10 μm,属于B型淀粉粒;也发现大量C型淀粉粒,但未发现A型淀粉粒。说明在淀粉体排布上,谷子与小麦也存在一定差异,这也是造成小米适口性较差的原因之一。在谷子品质改良实践中,可改造淀粉合成基因、调控基因,增加谷子淀粉体体积、改变不同类型淀粉体的比例从而提升谷子品质。
目前,对于谷子品质形成的研究进展缓慢,而品质的成因主要由蛋白和淀粉决定,因此通过开展蛋白体结构和淀粉特征等表型研究,从而为从分子水平解析谷子品质的形成及影响因素奠定基础,也为从转基因及基因组编辑等方面改良谷子品质提供理论依据。
猜你喜欢
计算机应用与软件(2022年6期)2022-07-12
油气地质与采收率(2022年3期)2022-05-20
自然灾害学报(2022年2期)2022-05-10
纺织科学研究(2021年7期)2021-08-14
药学研究(2021年6期)2021-07-26
中国学校体育(2021年10期)2021-04-26
海峡科技与产业(2019年4期)2019-10-26
作文·初中版(2019年8期)2019-09-09
中国新技术新产品(2016年20期)2016-12-08
汽车维护与修理(2015年6期)2015-02-28
