
喀斯特山地草地土壤酶活性及土壤微生物碳代谢活性研究
2020-05-06 00:59金章利刘高鹏周明涛许文年
水土保持研究 2020年3期
金章利, 刘高鹏, 周明涛, 许文年
(1.湖北三峡职业技术学院, 湖北 宜昌 443000; 2.湖北金源绿通生态环境工程有限公司,湖北 宜昌 443000; 3.三峡大学 土木与建筑学院, 湖北 宜昌 443002)
对于局地生态而言,不仅仅植被是其中关键的组成部分,土壤也在这一生态循环中发挥着巨大的作用,能量的交换和循环都离不开土壤[1-2],这是植被赖以生存的基础条件之一;植被在生长过程中离不开必要的水分和养分条件[3],而土壤能够很好地涵养水源,同时能够将植被所需的氮、磷等营养元素源源地向植株输送,其养分及水分直接关乎植株生长发育,并对其群落分布产生直接的制约[4-5]。植被在其生长的过程中难以避免地会因四季变换等多种原因出现枯枝落叶等现象,这些枯枝落叶形成凋落物,微生物分解,进而转化为土壤养分等营养物质[5-6],从而利于植株根系吸收利用,在这一过程中土壤酶起着重要的催化作用,能够明显促进有机质分解[7],该酶在整个有机质转化过程中发挥着无可替代的作用,制约着整个土壤生态循环和能量交换。综合来看,植被和土壤之间不仅仅是依赖作用[8],更是互动效应,这使得二者在不断变化、相互影响。通过对草地生态开展大量研究发现[8-10],其植被类型的变化能够直接影响到凋落物状况,同时对微生物活动产生影响,进而对土壤产生的作用也有明显差异[11-12]。不仅仅是土壤碳通量,就是在整个陆地生态中,土壤呼吸也起着无可替代的作用,其微小的变化就会直接对碳平衡产生有效影响[13]。近些年来,随着生物分析技术的提升,对土壤养分及植被的相关研究不断增多,土壤及生物多样性之间的研究不断加深。
我国地域分布广阔,受经纬度分布的地理位置影响,其不仅呈现复杂的气候特点,而且造就了高低起伏连绵不断的地形地貌[14],除了平原、丘陵山地之外,在西南地区还广泛分布着生态脆弱的喀斯特地貌,就该地貌分布面积而言[15],我国西南地区分布最广,这种地形地貌高低起伏,植被分布较少,其生态性仅强于寒漠地区。因其独特的岩溶地貌特点,且具有典型的起伏分布,高山和峡谷交错分布[15-16],因而造就了典型的二元地貌,形成了典型的地上、地下地貌景观分布差异性,但二者又是密不可分的,受此地貌的制约和作用,其生物群落及植被分布也存在相当大差异,虽然该地区因为生态脆弱而分布的植被数量较少,但是因为地形差异显著而导致了生物资源分布的多样性[17],呈现明显差异的生境高度等,同时也分布着多种微生物。因海拔差异巨大,该区域多布峡谷地貌,在形成陡峭山坡的同时造就了崎岖的地表,加之风力及雨水侵蚀,形成了典型的岩溶地貌[14-15],由于该区域的土层稀薄,因此降雨难以充分在土壤中积蓄,故影响作物生长的水分供给;受制于脆弱的生态,该区域并无大面积较集中的植被分布,同时植株的高度等长势较差,生长速度明显缓慢,整个生态的抗逆性较差,且波动较大,一旦发生环境逆境[16-17],其自我修复能力难以充分发挥。加之近些年来该区域过度地开发利用,使得该区域生态环境更加脆弱,呈现典型的石头多、耕地少的问题,不仅土壤受侵蚀问题凸显,而且呈现出明显的石漠化[18],这种问题在短期内难以自我修复。脆弱的生态对当地居民日常生活的制约作用越来越明显,不仅影响了当地粮食作物的产量,还对局地生态产生了重要的制约,基于此,大量的学者从植被分布、土壤养分及耕作等角度来探究该地区的地形地貌特点,本文将在结合以往研究成果的前提下,通过连续三年的实地研究来分析该区域的土壤特点及地形地貌,从而分析该地形地貌之下生物多样性及其养分特点,为喀斯特生态保护提供有益的参考和借鉴。
1 材料与方法
1.1 研究区概况
该试验区域为贵州省晴隆县境内,喀斯特地貌占据了该区域50%以上的面积,且高原峡谷密布,因海拔差异巨大,该区域多布峡谷地貌,在形成陡峭山坡的同时造就了崎岖的地表,加之风力及雨水侵蚀,形成了典型的岩溶地貌;受制于脆弱的生态,该区域并无大面积较集中的植被分布,同时植株的高度等长势较差,生长速度明显缓慢,整个生态的抗逆性较差,且波动较大,一旦发生环境逆境,其自我修复能力难以充分发挥。因其所处经纬度的影响,该区域的亚热带季风气候特点显著,且雨热同期,受地形地貌的影响,其不同的海拔之处具有较大差异的气候特点。试验区域的年均降雨量接近1 700 mm,年平均温度约为15℃,累积日照达1 500 h,全年无霜期超过300 d;该区域全年整体来讲具有较为充足的降雨和光照条件,但是由于该区域的土层稀薄,因此降雨难以充分在土壤中积蓄(图1),故影响作物生长的水分供给,从而形成了该区域以山地草地、乔木为主的植被分布特点,同时因崎岖的地表及陡坡低谷而多发自然灾害[16-17]。

图1 研究区近3 a平均气温和降水量月值变化
1.2 试验设计
在研究过程中选择的植被为山地草地,本研究从2013年开始进行连续5 a的实地观测研究,选取的取样区域分别位于海拔690,1 020,1 330 m(表1),在各个取样区域随机设置50个样方,要求长、宽均为100 cm,然后对各个样方的物种及其长势特点进行计算并记录,以便于后续进行多样性分析。

表1 长期定位样点描述
1.3 土壤养分测定
在试验过程中需要进行土壤取样以分析土壤养分等特点,本研究采取四分法,首先取样土壤1 kg,并混合均匀,然后开始连续20 d的风干,之后开展土壤但、磷等相关养分含量的测定[19]。
1.4 土壤呼吸和酶活性测定方法
为了有效测定土壤呼吸,首先在研究区域内布设相应的标记监测点,并提前在土壤之中嵌入测试基座,并将其中的留存植被剪除。在测量过程中所选气体分析系统为美国Li-6400,同时借助于土壤呼吸室,对土壤呼吸的日常监测频率为间隔2 h,测量时间集中在6:00—18:00,为了尽可能降低试验误差,每次测定进行两次重复,同时借助于温度探针对土壤深处的温度进行测定,对于其周边10 cm内的土壤开展相应的湿度测定,本研究中借助TDR。为了进行土壤养分等相关指标,首先在监测点选取不同海拔的土样,将地表清理后至下10 cm深度内进行取样,混合均匀后带回实验室两份土样,其中一份进行连续20天风干处理,之后取出杂质后过筛,另一份保存以备后续进行酶活性测定。
对土壤微生物呼吸的测定借助于碱液吸收法,微生物代谢熵qCO2=MR/MBC[7-8]
水解酶测定:借助于对硝基苯酚法。
微生物群落指标测定:在利用Simpson指数的过程中为了避免出现负数,需要对原始数据进行放大1 000倍的处理,同时对相应数据进行长达72 h的温育处理,之后再开展相应的多样性对比分析。本研究通过AWCD来对微生物利用碳源的能力进行衡量,二者是明显的正向表示关系,其计算公式如下[9-10]:
AWCD=∑[(Ci-R)/31]
对于Ci,R而言,其代表的分别是第i孔及对照组吸光值,本研究中Ci-R不低于零。
H=-∑Pi(lnPi)
对于式中Pi而言,其主要是从相对吸光值的角度来分析第i孔的占比,其计算公式如下所示:
Pi=(Ci-Ri)/∑(Ci-Ri)
Ds=1-∑Pi
统计分析:在开展指标统计分析之前对数据进行相应的均值处理,并充分考虑标准误差,然后借助于Excel 2010整理相关数据,SPSS 21.0进行相关分析及显著性检验等,运用CANOCO 4.5探讨与环境因子之间的关系。
2 结果与分析
2.1 不同海拔山地草地土壤养分
通过对表2的分析不难发现,不同海拔的草地区域形成了较大差异的土壤pH值,整体而言,pH值最高的是中海拔区域土壤,其次是高海拔区域,而其值最低的是低海拔区域,通过相关分析得知,对于不同海拔的草地而言,其土壤pH之间的巨大差异也通过了0.05的显著性检验,此外,对于高海拔草地而言,其pH值存在的变化幅度较小,仅为0.82~0.83的区间内,而对于低海拔、中海拔而言,其草地土壤pH分别为0.76~0.79,0.87~0.91。受到海拔高度不同的制约草地土壤的全碳含量也呈现了较大差异,其中含量最高的是中海拔区域,其次是高海拔区域,而低海拔区域含量最少。通过相关分析得知,对于不同海拔的草地而言,其土壤全碳含量间的巨大差异也通过了0.05的显著性检验;此外,对于中海拔区域而言,其全碳变化幅度最小,仅为15.48~15.67 g/kg,而低海拔、高海拔区域的全碳含量变化区间分别为12.36~13.25,13.56~13.89 g/kg。从全氮的角度来看,海拔仍然对其含量产生了较大的制约效果,通过分析得知,其含量最高的是中海拔区域,其次是高海拔,而低海拔区域含量最少,通过相关分析得知,对于不同海拔的草地而言,其全氮含量间的巨大差异也通过了0.05的显著性检验,这与全碳含量的变化基本一致;此外,对于低海拔和高海拔区域而言,其全氮变化幅度最小,分别为1.16~1.17,1.22~1.23 g/kg,而中海拔区域的全氮含量变化区间处于1.28~1.30 g/kg。从全磷的角度来看,通过对比发现,其含量最高的是中海拔区域,其次是高海拔,而低海拔区域含量最少,通过相关分析得知,对于不同海拔的草地而言,其全磷含量间的微小差异并未通过0.05的显著性检验,这也是其与全碳、全磷等方面的较大差异。从全钾、碱解氮和速效磷的角度来看,通过对比发现,其呈现基本一致的变化规律,也就是说其值最高的是中海拔区域,其次是高海拔区域,而含量最低的是低海拔区域,其含量之间的差异通过了0.05的显著性检验。

表2 不同海拔山地草地土壤养分效应
2.2 不同海拔山地草地土壤微生物对碳源利用强度
对于微生物而言,其群落分布受到多种因素的制约,尤其是多种影响因素的交互作用影响更为复杂,其多样性分布虽然能够对群落总体变化进行有效的体现,但是无法充分反映其新陈代谢机理,为了探究海拔因素对于微生物碳源利用水平的影响,本研究借助于AWCD值分析来探究微生物多样性,这是对其光密度加以充分考虑,由于其化学基团具有较大的性质差异,从而对碳源进行不同的分类处理,本研究中共进行聚合物、胺类等6种处理。通过对表3的分析不难发现,由于碳源特性的差异,也导致了微生物对其利用水平有所差异,但是整体而言,这种差异较小,通过对其连续三年的观测值求均值得知,其利用率最高的是中海拔区域,其次是高海拔区域,而低海拔的利用率最低,其中对于氨基酸类而言,受海拔差异的影响,其微生物碳源利用率虽然有所差异,但是并未通过0.05的显著性检验,但是从羧酸类、聚合物类的角度来看,中海拔区域具有更高的含量,其差异达到了0.05的显著性水平,对于胺类来说亦是如此。总体而言,通过连续三年的观测研究得知,随着时间的推移,这6种碳源物质呈现上升趋势,其中微生物对碳水化合物、羧酸类的利用水平更高,而胺类最低。
2.3 不同海拔山地草地土壤微生物群落多样性
为了探究微生物对碳源的利用情况,本研究借助于其群落分布的多样性来加以对比分析,也就是通过AWCD值来计算其多种指数,尤其是物种丰度H、均匀度E,此外还要探究其优势度Ds、碳源利用丰度S(表4)。总体而言,通过连续三年的观测研究得知,随着时间的推移,对于微生物多样性指数而言,其上升趋势较为明显,尤其是中海拔区域,其次是高海拔,整体来说,中海拔区域更高,其差异具有显著性(p<0.05)。

表3 不同海拔山地草地土壤微生物对碳源利用强度
注:同列不同小写字母表示差异显著(p<0.05)

表4 不同海拔山地草地土壤微生物群落多样性
2.4 不同海拔山地草地土壤呼吸及微生物代谢熵
表5为不同海拔山地草地土壤呼吸及微生物代谢熵,不同海拔山地草地土壤微生量碳、微生物呼吸和微生物代谢熵2015—2017年均呈增加趋势;3 a内土壤微生物呼吸平均值基本表现为中海拔>高海拔>低海拔,并且不同海拔差异均显著(p<0.05);3 a内土壤微生物量碳和土壤微生物代谢熵平均值基本表现为中海拔>高海拔>低海拔,低海拔和高海拔土壤微生物量碳和土壤微生物代谢熵差异不显著(p>0.05),二者显著低于中海拔(p<0.05)。
2.5 不同海拔山地草地土壤酶活性
表6表明不同海拔山地草地土壤酶活性有一定的差异,其中不同海拔山地草地土壤糖苷酶、几丁质酶、亮氨酸氨基肽酶、碱性磷酸酶、酚氧化酶δ和过氧化物酶δ从2015-2017年均呈增加趋势;3 a内土壤糖苷酶、几丁质酶、亮氨酸氨基肽酶、碱性磷酸酶、酚氧化酶δ平均值基本表现为中海拔>高海拔>低海拔,不同海拔土壤糖苷酶、几丁质酶、亮氨酸氨基肽酶、碱性磷酸酶、酚氧化酶δ差异均显著(p<0.05);过氧化物酶δ平均值基本表现为中海拔>高海拔>低海拔,不同海拔土壤过氧化物酶δ差异均不显著(p>0.05)。

表5 不同海拔山地草地土壤呼吸及微生物代谢熵
注:同列不同小写字母表示差异显著(p<0.05)。
2.6 土壤酶活性与微生物群落多样性之间相关性
为了探究酶活性对微生物的作用机理,本研究借助于RDA排序法进行分析对比(图2),通过研究得知,从第1排序轴而言,其排序较为突出的不仅包括微生物呼吸,还包括其代谢熵,二者的箭头连线最长,不仅与物种第1排序轴之间的正向变化关系通过了0.01的检验水平,而且与环境因子通过了0.01的显著性检验,这种较为重要的位置说明了二者能够显著影响微生物分布,尤其是其中土壤几丁质酶。对于微生物生长发育及新陈代谢而言,其受到水分、土壤等多种因子的制约,在本研究过程中借助于冗余分析RDA来加以探究,探究的主要解释变量不仅包括土壤养分,还包括其酶活性,这样能够更为直观地对比分析影响因子的作用效果,形成更为鲜明的对比。从中不难看出,对于前两个排序轴而言,其拥有高达89%的累积解释度,其中解释率最高的是第一个因子,同时其影响效果通过了0.05的显著性检验,说明其制约效果最强。另外,微生物丰度对于土壤养分之间的正向影响效果较为突出,土壤养分的增强对于促进微生物多样性效果显著,尤其是全碳、全氮含量的提升。

表6 不同海拔山地草地土壤土壤酶活性
注:同列不同小写字母表示差异显著(p<0.05)。
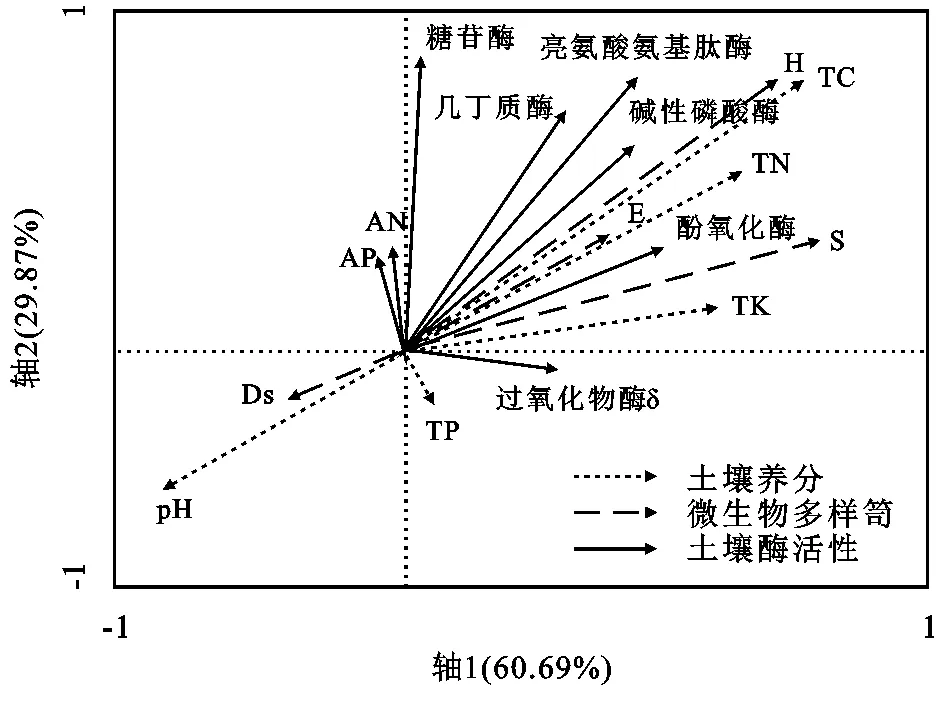
图2 土壤酶活性与微生物多样性之间RDA排序
3 讨论与结论
作为本区域主要的植被分布,山地草地群落分布特点受到环境的多方面影响,其在生长过程中离不开必要的水分和养分条件[20],而土壤能够很好地涵养水源,同时能够将植被所需的氮、磷等营养元素源源地向植株输送[21],其养分及水分直接关乎植株生长发育,并对其群落分布产生直接的制约,因此来说,只有充分深入分析植被、土壤等环境因子之间的关系才能为保护生态奠定基础。通过发达的根系,植株能够及时进行养分传输,其所需的养分能够从土壤中获取,之后再借助于其根系输送至植株各部分,这样能够将土壤养分有效供给植株,从而满足其生长发育,这样在根系的作用下,植物实现了与土壤等外界环境的能量交换[25]。其根系发达程度越强,说明其对土壤养分吸收利用的能力越强,这样其对土壤养分的利用效能越高;此外,根系生物量的分布受根系分布的影响。王启基等[24]通过实地研究分析得知,植被群落生产力受到土壤养分的直接作用,二者呈现典型的正向线性关系。通过本研究分析得知,受海拔的影响,土壤所含的全碳、全氮、全钾含量出现明显的差异,且在0.05检验水平下达到显著,且中海拔区域的含量最高[22-23],其次是高海拔,而低海拔区域最低,碱解氮和速效磷的变化亦是如此;而土壤pH值的变化则与之相反;全磷含量的变化并不受海拔的显著影响,也即是并未在0.05检验水平下显著。通过本研究实地调查得知,随着海拔的上升,土壤养分在中海拔区域达到最大,且该区域具有明显的植被分布多样性,而高海拔区域由于温度相对较低,其土壤养分呈现下降趋势,进而影响群落分布及生物量,整体来看,三者随着海拔的升高而先升后降;磷含量最主要的影响因素是成土母质,因此与海拔的差异联系不大[24]。在海拔上升的过程中,其水热条件相对上升,凋落物增加等导致有机质含量有所提高,微生物能够获取更多的能量,其代谢熵及呼吸增强,进而使得其活性提升,能够加强对土壤养分的吸收利用。
本研究中通过AWCD来反映微生物活性,这代表着其对碳源的利用水平,同时也能够对其群落分布特点进行充分反映[13-14],对于AWCD指标而言,其值越大说明微生物分布密度更大,同时也说明其具有较强的活性,其值越小说明其活性越弱。通过对比分析不同海拔微生物的状况得知,就微生物对6种碳源利用水平而言,中海拔区域的微生物具有更强的碳源利用效能,其次是高海拔区域,而低海拔区域利用能力较弱,但是氨基酸类的吸收利用能力并不受海拔的显著影响,也就是说在0.05检验水平下并不显著,对于羧酸类、聚合物类和胺类而言,其在低海拔及高海拔之间的差异并不显著,同时二者均未超过中海拔区域。综合来看,经过连续三年的试验观测,虽然海拔不同,但是本试验中的6种碳源均是逐年上升的走势,其中微生物利用能力最强的碳源是碳水化合物和羧酸类,而利用效能最低的碳源是胺类。
对于微生物而言,不仅其生物量碳受到环境的明显制约,同时其呼吸也随外部环境的变化而变化,这也是其碳代谢特征的两大要素,能够较有效地对生态环境进行指示,能够有效作为土壤生态的重要反映指标,及时反映其变化性[25-26]。作为土壤有机质的重要组成部分,生物量碳具有较高的活性,成为活性有机碳的重要来源之一,同时其周转速度较快,外部环境轻微的变化即能引起其活性等方面的变化,因此能够及时反映出土壤的细微变化,进而对土壤碳库加以反映[21-22]。对于微生物呼吸而言,其呼吸过程也是对有机质的分解和降解过程,是对有机质进行潜在的矿化过程,体现的是其整体活性及代谢水平,其变化状况反映着外部环境的变化,其变化方向是对环境胁迫的响应[22]。微生物代谢熵能够对土壤两个方面加以充分体现。首先,该指标体现了微生物群落分布状况,是其对能量维持的结果;其次,能够对其利用有机质的效能加以反映;这样,微生物量碳及其活性能够充分结合起来,进而通过对其连续的观测研究来分析外部环境变化,因此来说,该指标是微生物活性的重要体现指标之一[27-28]。微生物在新陈代谢过程中会对碳源等进行一定的消耗,其代谢熵值较大,说明其在呼吸过程中消耗了较大比例的碳,那么在这种情况下,能够用于构筑其细胞的碳就会相应减少,这样微生物就难以获得较高利用率的碳,不利于土壤质量的稳定[29-30],因此说较小的代谢熵有利于维持土壤质量。通过连续三年的对比分析得知,虽然海拔存在较大差异,但是其区域内的土壤酶活性均呈现上升趋势,且不同海拔间的土壤酶活性存在较大差异,其中最大值出现在中海拔区域,其次是高海拔,而最低的是低海拔区域。
在海拔不断上升的情况下,出于温度及水分条件的制约,不仅植被凋落物数量明显下降,而且有机质含量明显减少,而微生物的呼吸所需能量主要来源之一就是有机质,因此会明显抑制其呼吸作用,同时降低微生物量碳,最终导致微生物难以充分进行新陈代谢,甚至影响其群落分布,大大降低其活性;微生物在进行有机质分解及降解过程中,难以避免地需要消耗一定的能量来维持其新陈代谢活动,而其获取能量的水平受损,最终影响了其对土壤养分的利用。中海拔土壤含有丰富的有机物质,为土壤微生物生长提供更多碳源和可利用养分,促进土壤微生物的大量繁殖以及微生物的活动,导致土壤微生物对有机质的利用效率和矿化速率增加,在形成更多微生物生物量碳的同时释放CO2,进而使土壤微生物代谢熵降低[26-27]。因此来说,对于中海拔区域,山地草地的土壤微生物能够较充分地进行有机质分解,利于土壤养分保持。通过对研究数据进行相关分析得知,对于土壤酶活性而言,其受到微生物群落分布的影响,二者存在显著的相关性;通过相关系数对比可以看出,酶活性对优势度的影响并不大;通过RDA排序可知,微生物呼吸及代谢熵显著影响其多样性分布。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
散文诗(2021年22期)2022-01-12
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
幼儿100(2020年31期)2020-11-18
天津科技(2020年4期)2020-05-09
疯狂英语·初中版(2019年4期)2019-09-10
环球时报(2019-05-23)2019-05-23
小太阳画报(2018年6期)2018-05-14
郑州大学学报(工学版)(2014年6期)2014-03-01
