
饶河地区野生动物携带病毒的基因分型及序列特征分析
2020-05-05 02:37王奥男党蕴琦李博琦李永久
中国人兽共患病学报 2020年3期
王奥男,党蕴琦,李博琦,李永久,刘 铸
黑龙江省饶河县地处中俄边境,是中俄贸易重要口岸,该地区包含湿地沼泽和山地森林,物种多样性丰富。随着新发和复发传染病给全球带来越来越重的疾病负担,人们开始注重决定疾病风险和相关生态学过程的环境因素的问题[1-2]。很多传染病的暴发和人类的贸易、鸟类的迁徙等息息相关。虫媒传染病依靠吸血节肢动物进行传播,是许多自然疫源性疾病的病原体。对环境变化极为敏感,其错综复杂的生态过程影响病媒在环境中的分布和数量,以及其与人类和非人类贮主宿主间的接触[3-5]。很多自然疫源性疾病的病原体(包括虫媒病毒)储存于小型哺乳动物,特别是啮齿动物体内,因此啮齿动物对自然疫源性疾病的传播具有重要作用[6]。另外,鸟类是自然界中病原体的重要贮存宿主。
黄病毒属病毒(Flavivirus)和甲病毒属病毒(Arenavirusgenus)属于虫媒病毒,其中大多数可自然感染多种脊椎动物,主要是鸟类、鼠类。饶河地区是候鸟迁徙的重要迁徙地和栖息地,鼠类资源丰富。黑龙江省是最严重、最典型的蜱传脑炎病毒(Tick-borne encephalitis virus)分布流行区之一[7]。汉坦病毒(Hantavirus, HV)属于布尼亚病毒科(Bunyaviridae)汉坦病毒属(Hantavirus),自然宿主为啮齿类动物。HV对啮齿类只造成一过性感染,对宿主的寿命和繁衍并无影响,但其可通过宿主的排泄物感染人类并引发两种严重的临床疾病:肾综合征出血热(hemorrhgic fever with renal syndrome, HFRS)和汉坦病毒肺综合症(hantavirus cardiopulmonary syndrome, HPS)。我国存在汉城型(Seoul virus,SEOV)和汉滩型(Hantaan virus,HTNV)两种 HV 型别,中国拥有世界上最高的HFRS发病率,而黑龙江是最严重的疫区之一[8]。
本研究对饶河地区部分野生动物进行黄病毒属病毒、甲病毒属病毒和汉坦病毒等病原体筛查,对检测出的阳性样本的基因片段进行分型和核苷酸序列分析,确定当前饶河地区宿主动物所携带病原体的基因型别,分析其变异情况,对该地区自然宿主动物的感染情况进行监测。
1 材料与方法
1.1样本的采集与处理 本次样本由饶河检疫局口岸科提供(2018年秋季野外捕捉),将样本分类鉴定(表1),鉴定完毕后无菌解剖,取完整脏器(脑、肺、肝、脾、肾、膀胱、心、肠、胃)分别放入冻存管内,置于-80 ℃超低温冰箱内保存。
表1 待测样本的物种和数量
Tab.1 Species and number of samples to be tested

物 种数量东方田鼠(Microtus fortis)9褐家鼠(Rattus norvegicus)5黑线姬鼠(Apodemus agrarius)5黄鼬(Mustela sibirica)1黑龙江刺猬(Erinaceus amurensis)1绿头鸭(Anas platyrhynchos)2白骨顶鸡(Fulica atra)1松鼠(Sciurus vulgaris)52总 计76
1.2RNA的提取 将每只动物的每个脏器分别采用TRIZOL(Takara)法提取RNA。RNA溶解于30 μL RNase-free water。
1.3引物设计与合成 本次研究所需引物(表2)由北京华大基因合成。
表2 引物详细信息
Tab.2 Primer detailed information

引物序列(5′-3′)位置/bp黄病毒属病毒通用引物[9](Flavivirus)HF: AATGTATGCAGATGATACAGCAGGNTGGGAYACHR: TCTATCATCAATTGGTTTAACAACACARTCRTCNCC421-445 甲病毒属病毒通用引物[10](Arenavirusge-nus)JF: GGAGTTAGAACGGCGTATTGGATAGGNTTYGAYACJR: CATCTGTAGCCAGGATGCCTGTCATYTGRTCRC510-550 巢式PCR第一轮汉坦病毒通用引物[11](HV)HVF: AAAAGTAGGTGITAYATCYTIACAATGTGGHVR: GTACAICCTGTRCCIACCCC445 巢式PCR第二轮汉坦病毒汉城型引物[11](SEOV)SEOVF: GTGGACTCTTCTTCTCATTATTSEOVR: TGGGCAATCTGGGGGGTTGCATG418 巢式PCR第二轮汉坦病毒汉滩型引物[11](HTNV)HTNVF: GAATCGATACTGTGGGCTGCAAGTGCHTNVR: GGATTAGAACCCCAGCTCGTCTC383
1.4RT-PCR扩增(TAKARA RR057A试剂盒) 将RNA产物各取10 μL,结合表1,按物种和器官类型进行混合,如东方田鼠脑(90 μL)、东方田鼠肝(90 μL)、褐家鼠肺(50 μL)。反应条件如下:Prime Script 1 step Enzyme Mix 1 μL,2×1 step Buffer(Dye Plus)12.5 μL,RNase Free dH2O 8.5 μL,上游引物 1 μL,下游引物 1 μL,RNA混合模板 1 μL,合计25 μL;50 ℃ 30 min,94 ℃ 2 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min(返回第3步,35个循环),4 ℃ ∞。
1.5PCR产物鉴定及核苷酸序列测定 取扩增产物5 μL进行琼脂糖凝胶电泳鉴定。若条带的相对分子质量与预期片段大小相同,则表明为特异性扩增产物。扩增产物由北京市华大基因公司进行核苷酸序列测序。
1.6核苷酸序列分析 应用DNAStar软件包对核苷酸序列进行同源性比较,构建系统进化树和氨基酸序列的比对,其中引用序列均来自GenBank。
2 结 果
2.1PCR扩增产物电泳鉴定及测序鉴定 经电泳鉴定,所有样本中均无黄病毒属和甲病毒属的目的片段。S11 (褐家鼠肺)、S13 (褐家鼠肺)、S14 (褐家鼠肺)、SS7 (松鼠肺)合计4个样本中疑似携带HV目的片段(图1)。将疑似阳性样本PCR产物交送北京华大基因进行核苷酸序列测序,结果显示S11,S13,S14,SS7携带SEOV。
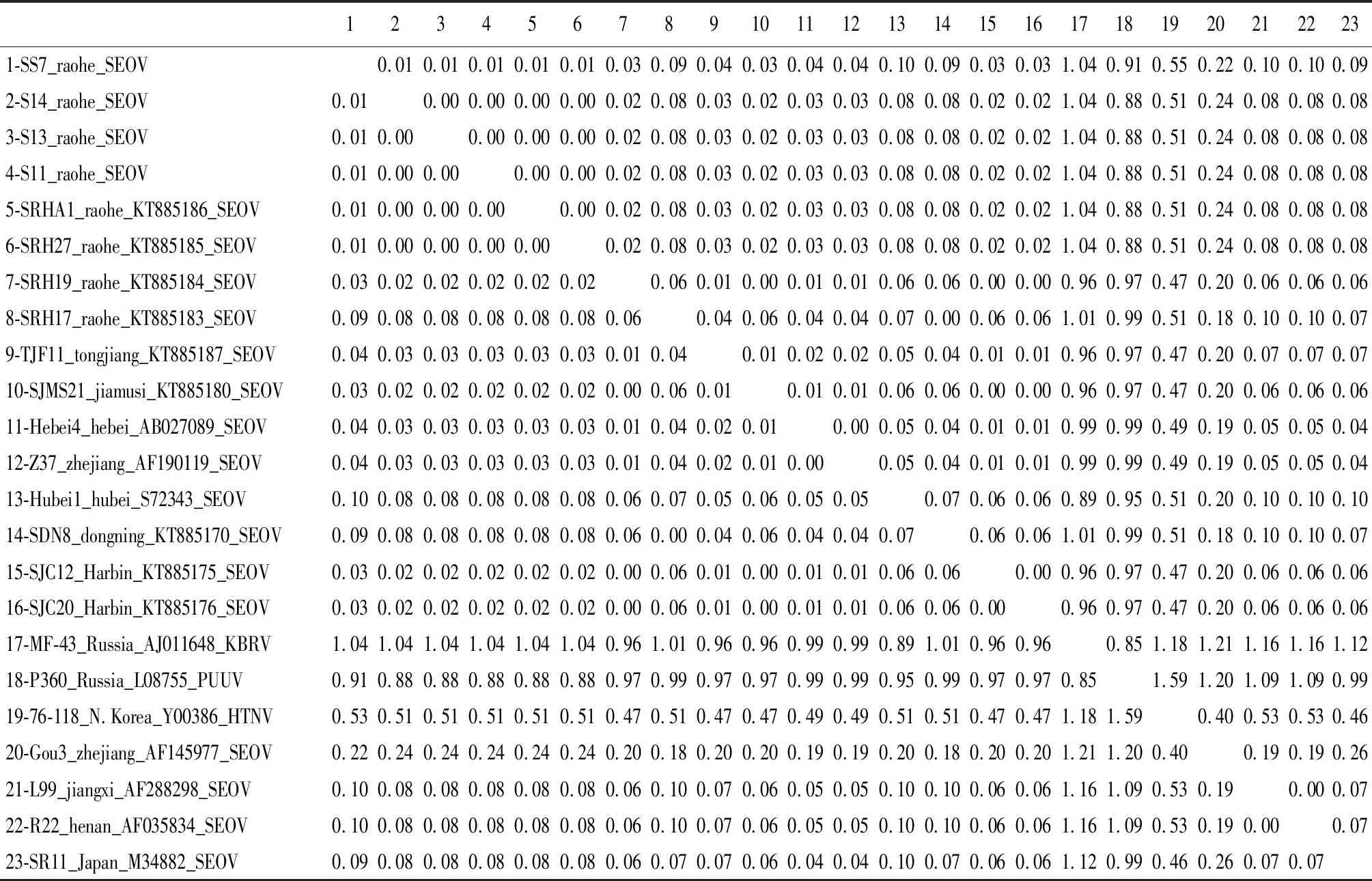
2.2SEOV分离株的M基因部分片段遗传多样性分析 以本次饶河地区分离出的4株SEOV及来自GenBank的19株国内外标准毒株进行核苷酸序列比对和同源性比较(表3)。结果显示,本次饶河地区3只褐家鼠S11、S13、S14和1只松鼠SS7中分离出的SEOV之间遗传距离范围为0.00~0.01,相互之间同源性很高。与黑龙江地区其他SEOV病毒株之间的遗传距离也相对很近。
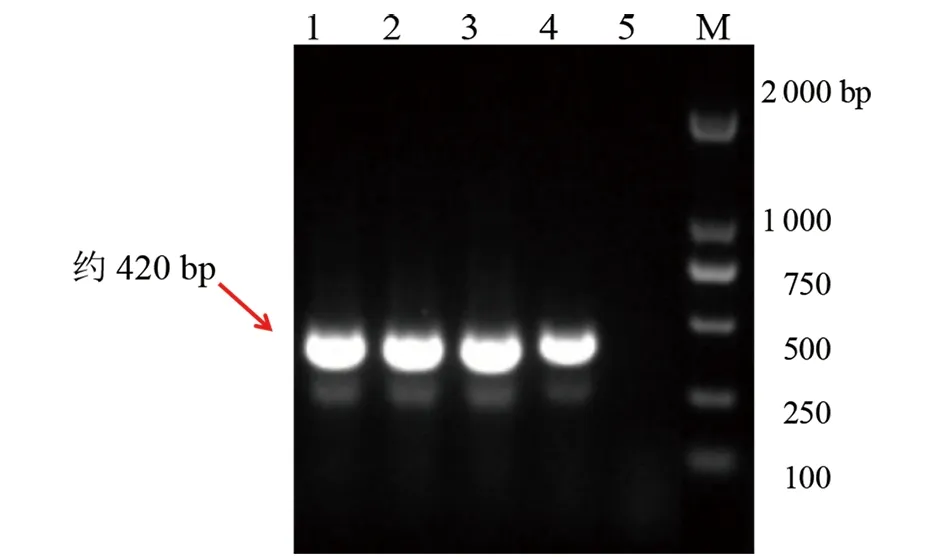
注:1-4模板均为肺部PCR产物,1-3为褐家鼠,4为松鼠,5为阴性对照,6为2 000 bp Marker图1 汉坦病毒初筛套式PCR产物电泳结果Fig.1 SEOV PCR product electrophoresis results
2.3SEOV分离株与HV原型毒株的系统进化分析 以本次饶河地区分离出的4株SEOV的M基因部分片段与国内外有代表性的19株SEOV的M基因部分片段共同构建系统发生树(图2)。通过系统进化分析发现S11、S13、S14携带的SEO型HV亲缘关系最近,SS7稍远,但总体亲缘关系十分相近。与此前黑龙江省哈尔滨、佳木斯、饶河、同江,以及河北、浙江的SEOV病毒株序列亲缘关系相对较近。与日本、江西、河南的SEOV病毒株序列亲缘关系相对较远。
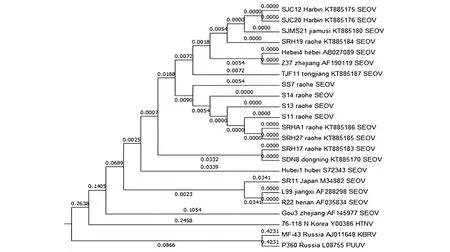
图2 多种病毒M片段核苷酸序列系统进化树Fig.2 Phylogenetic tree of nucleotide sequences of various viral M fragmentsresults
表3 多种病毒M片段核苷酸序列同源性比较
Tab.3 Comparison of nucleotide sequence homology of various viral M fragments
12345678910111213141516171819202122231-SS7_raohe_SEOV0.010.010.010.010.010.030.090.040.030.040.040.100.090.030.031.040.910.550.220.100.100.092-S14_raohe_SEOV0.010.000.000.000.000.020.080.030.020.030.030.080.080.020.021.040.880.510.240.080.080.083-S13_raohe_SEOV0.010.000.000.000.000.020.080.030.020.030.030.080.080.020.021.040.880.510.240.080.080.084-S11_raohe_SEOV0.010.000.000.000.000.020.080.030.020.030.030.080.080.020.021.040.880.510.240.080.080.085-SRHA1_raohe_KT885186_SEOV0.010.000.000.000.000.020.080.030.020.030.030.080.080.020.021.040.880.510.240.080.080.086-SRH27_raohe_KT885185_SEOV0.010.000.000.000.000.020.080.030.020.030.030.080.080.020.021.040.880.510.240.080.080.087-SRH19_raohe_KT885184_SEOV0.030.020.020.020.020.020.060.010.000.010.010.060.060.000.000.960.970.470.200.060.060.068-SRH17_raohe_KT885183_SEOV0.090.080.080.080.080.080.060.040.060.040.040.070.000.060.061.010.990.510.180.100.100.079-TJF11_tongjiang_KT885187_SEOV0.040.030.030.030.030.030.010.040.010.020.020.050.040.010.010.960.970.470.200.070.070.0710-SJMS21_jiamusi_KT885180_SEOV0.030.020.020.020.020.020.000.060.010.010.010.060.060.000.000.960.970.470.200.060.060.0611-Hebei4_hebei_AB027089_SEOV0.040.030.030.030.030.030.010.040.020.010.000.050.040.010.010.990.990.490.190.050.050.0412-Z37_zhejiang_AF190119_SEOV0.040.030.030.030.030.030.010.040.020.010.000.050.040.010.010.990.990.490.190.050.050.0413-Hubei1_hubei_S72343_SEOV0.100.080.080.080.080.080.060.070.050.060.050.050.070.060.060.890.950.510.200.100.100.1014-SDN8_dongning_KT885170_SEOV0.090.080.080.080.080.080.060.000.040.060.040.040.070.060.061.010.990.510.180.100.100.0715-SJC12_Harbin_KT885175_SEOV0.030.020.020.020.020.020.000.060.010.000.010.010.060.060.000.960.970.470.200.060.060.0616-SJC20_Harbin_KT885176_SEOV0.030.020.020.020.020.020.000.060.010.000.010.010.060.060.000.960.970.470.200.060.060.0617-MF-43_Russia_AJ011648_KBRV1.041.041.041.041.041.040.961.010.960.960.990.990.891.010.960.960.851.181.211.161.161.1218-P360_Russia_L08755_PUUV0.910.880.880.880.880.880.970.990.970.970.990.990.950.990.970.970.851.591.201.091.090.9919-76-118_N.Korea_Y00386_HTNV0.530.510.510.510.510.510.470.510.470.470.490.490.510.510.470.471.181.590.400.530.530.4620-Gou3_zhejiang_AF145977_SEOV0.220.240.240.240.240.240.200.180.200.200.190.190.200.180.200.201.211.200.400.190.190.2621-L99_jiangxi_AF288298_SEOV0.100.080.080.080.080.080.060.100.070.060.050.050.100.100.060.061.161.090.530.190.000.0722-R22_henan_AF035834_SEOV0.100.080.080.080.080.080.060.100.070.060.050.050.100.100.060.061.161.090.530.190.000.0723-SR11_Japan_M34882_SEOV0.090.080.080.080.080.080.060.070.070.060.040.040.100.070.060.061.120.990.460.260.070.07
3 讨 论
HV为分节段的单股负链RNA病毒,病毒的基因组分大(L)、中(M)、小(S)3个片段,3个基因片段的核酸变异以M基因片段的变异最显著,M片段编码病毒糖蛋白G1和G2的前体蛋白,M基因片段的变异会导致HV在细胞融合、结合病毒受体、血凝活性、包膜糖蛋白的表达以及糖基化等方面发生改变,从而影响HV的致病性[12]。20世纪80年代初期,我国首次分离出HV,此后通过病原学证实我国存在SEOV和HTNV两种HV型别[13]。褐家鼠为SEOV原始宿主,黑线姬鼠为HTNV原始宿主[14-15]。黑龙江地区是SEOV和HTNV的混合型疫区[16],是我国最早发生HFRS的地区,发现初期属于以姬鼠型为主的混合型疫区,却已有向混合型演变的趋势[17]。本次研究发现饶河地区鼠类携带的HV以SEOV为主;亲缘关系与黑龙江省其它地区常见的SEOV较近;除褐家鼠外,松鼠也携带SEOV。因本次实验样本量数量有限,虽未检测出HTNV、黄病毒属病毒和甲病毒属病毒,但不能排除该型病毒在本地区宿主动物间的存在。许多疫区都存在着各自的优势HV基因型或亚型,这可能是因HV与本疫区的优势自然宿主共进化造成的[18]。
分子流行病学表明,目前所有SEOV毒株在基于M基因部分片段的系统发生树上可分为6个分支[19-20]。进化分支1、3和4中包含的毒株主要集中在中国东北[21-22]。本次饶河地区只分离到了属于进化分支3的HV毒株[21,23-24]。从基因系统发生树上来看,SS7毒株与SEOV序列十分相似,并且从系统发生树上可推断出SS7毒株属于SEOV的远东地区病毒进化分支。
黑龙江地区的气候和自然地理条件非常适合携带HV宿主动物的生存和繁殖,但是随着人们生活条件的改善,禁止伐木狩猎,家鼠和野鼠与人类交集的逐渐减少,HFRS在黑龙江省暴发的概率也越来越小。但褐家鼠有着强大的生存适应力,在全球范围都十分活跃,与人类活动关系密切,并且喜欢栖居于建筑物内。饶河作为中俄贸易重要口岸,更要做好疫源监测和预防宣传工作,防止传染病随着进出口贸易而广泛传播。
利益冲突:无
猜你喜欢
动物医学进展(2022年9期)2022-11-26
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
中国饲料(2019年19期)2019-03-25
小星星·阅读100分(高年级)(2015年11期)2015-11-28
