
两个玉米品种在萌芽期和苗期的干旱耐性比较分析
2020-04-29 03:16:05李玉华范春丽雷志华张丽萌卢园园王同朝
西北大学学报(自然科学版) 2020年5期
李玉华 范春丽 雷志华 张丽萌 卢园园 王同朝

摘要:以登海605(DH)和偉科702(WK)2个抗旱性不同的玉米品种为材料,分别在萌芽期采用精确称重控水,在苗期采用20%PEG-600溶液模拟田间土壤干旱胁迫的方法,对2个玉米品种萌芽期的生长量特征参数,苗期叶片光合作用特征参数、叶绿素荧光及多项生理指标进行测定,比较研究干旱胁迫对这2个玉米品种种子萌发和幼苗生理性状的影响。研究结果表明,在干旱胁迫条件下,2个玉米品种在萌芽期的发芽率、发芽势、发芽指数、种子萌发抗旱指数、株高、胚根长、贮藏物质转运率、地上和地下部分干重(与对照相比)均下降,且DH各指标降低的幅度均低于WK下降的幅度;干旱胁迫使2个玉米品种苗期叶片的净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)均降低,且DH的Pn,Tr和Gs下降幅度均低于WK,但对2个品种玉米叶片的荧光参数几乎没影响;干旱处理使2个玉米品种叶片的过氧化氢(H2O2)、丙二醛(MDA)和质膜透性均升高,且DH叶片中H2O2,MDA含量和质膜透性升高幅度均小于WK,同时,干旱处理使2个玉米品种叶片的抗氧化酶CAT和POD活性、脯氨酸、还原型抗坏血酸(AsA)、还原型谷胱甘肽(GSH)及可溶性蛋白含量均升高,DH叶片中这些指标升高幅度均较大。综上分析得出,干旱胁迫主要抑制2个玉米品种叶片的光合作用和碳水代谢速率,进而抑制其生长,二者均通过提高保护酶SOD,POD, CAT和APX活性,增加渗透调节物质等策略来缓解干旱胁迫造成的伤害。DH多项参数比WK受干旱影响小,较耐旱,研究结果为农业生产选育抗旱玉米品种提供了一定的理论依据。
关键词:玉米;干旱胁迫;生长特征;光合特性;抗氧化酶
中图分类号:S513
DOI:10.16152/j.cnki.xdxbzr.2020-05-002 开放科学(资源服务)标识码(OSID):
Comparative analysis on drought tolerance of two maize
cultivars at germination and seedling stages
LI Yuhua1,2, FAN Chunli1, LEI Zhihua1, ZHANG Limeng1,LU Yuanyuan1, WANG Tongchao2
(1.School of Life Sciences, Zhengzhou Normal University, Zhengzhou 450044, China;
2.College of Agronomy, Collaborative Innovation Center of Henan Grain Crops, Zhengzhou 450002, China)
Abstract: Two maize cultivars of Denghai605 (DH) and Weike702 (WK) with different drought resistance were planted in an artificially controlled water environment during the germination stage, and the 20% PEG-600 solution during the seedling stage were used as materials to study the effects of drought stress on their growth parameters, the photosynthetic and physiological characteristics as well as other physiological indicators at germination and seedling stage. The results showed that, under drought stress, the germination rate, germination potential, germination index, germination drought resistance index of seed, plant height, radical length, root volume, storage matter transformation rate and the dry weight of aboveground and underground (compared with the control group) of the two maize cultivars all decreased at the germination stage, and the decrease of each index of DH was lower than that of WK; drought stress reduced the net photosynthetic rate (Pn), transpiration rate (Tr) and stomatal conductance (Gs) of the leaves of the two maize cultivars, and the decrease of Pn, Tr and Gs of DH was lower than that of WK. However, little effect was exerted on the fluorescence parameters of the leaves of two maize cultivars; drought stress increased the contents of H2O2, MDA and plasma membrane permeability in leaves of two maize cultivars seedlings, and the increase of indexes in DH leaves were less than that of WK; in addition, the activities of antioxidant enzymes CAT and POD, proline, reduced ascorbic acid, reduced glutathione and soluble protein in leaves of two maize cultivars were increased by drought stress, and these indicators of DH increased much more than that of WK. To sum up, drought stress mainly inhibits photosynthesis and carbohydrate metabolism rate of maize leaves and further inhibits their growth, both of which alleviate the damage of drought stress by improving the activities of protective enzymes SOD, CAT, POD and APX and increasing osmotic adjustment substances. The physiological parameters of DH were less affected by drought than that of WK, DH is more drought-resistant than WK. The findings provide certain theoretical basis for breeding drought-resistant maize cultivars in agricultural production.
Key words: maize; drought stress; growth characteristics; photosynthetic characteristics; antioxidant enzyme
干旱胁迫对植物的结构、代谢和生长有一定的影响,通常表现为细胞膨压降低、气孔闭合、水分损失、叶片水势降低、细胞伸长和生长缓慢等[1]。严重的水分缺乏会破坏光合作用、能量生产和新陈代谢,并最终导致细胞死亡[2]。叶片生长缓慢是水分供应有限的最早症状之一,这有利于植物减少叶片蒸腾和保水[3]。玉米(Zea mays L.)的播种面积和总产量均位居全世界第三位,仅次于水稻和小麦,在我国的种植面积和产量又是位居秋粮作物首位。玉米在其全生育期内需水量大,是一种对低水分胁迫敏感的作物,萌发期和苗期对干旱胁迫十分敏感,若在生殖生长阶段,如抽雄期、开花期、抽丝期等遭遇持续干旱,将会严重降低玉米产量甚至绝收。据研究,由于干旱造成的玉米减产,超过其他因素造成的减产总和[4]。研究玉米萌发出苗及苗期的抗旱性机制,是鉴定和选育玉米抗旱品种的前提。因此,筛选玉米抗旱种质资源,并培育抗旱节水品种对玉米生产具有重要意义。
前人已經选用不同玉米品种(自交系和杂交种),不同发育时期,不同程度的干旱胁迫及控制不同的胁迫时间等方式方法,对玉米抗旱农艺性状、生理生化、分子响应机制及对产量影响做了大量的研究工作[5-8],确定了多项玉米抗旱性鉴定的指标[9],筛选出了一批抗旱种质资源[10-11],并获得了大量响应干旱胁迫的基因和基因产物[12-14]。研究发现,不同玉米品种在相同种植环境,同一玉米品种在不同种植环境,对干旱胁迫的适应性和抵御能力都是不相同的,它们在农艺性状、形态结构、生理生化及生长发育等方面,形成了复杂的适应和抵御干旱机制。
河南省近几年多次发生干旱,且旱情出现早,持续时间长,影响了玉米种子萌发和幼苗生长,给玉米产量造成很大损失。因此,有针对性地加强河南省玉米地方品种抗旱性研究是十分必要的。种子萌发期是决定玉米群体的重要阶段,与苗期关系密切,玉米种子活力高低直接决定出苗率高低及成株多少。在相同干旱环境下,若玉米品种出苗率高,整齐一致,且长势好,能反映出该品种种子有较高的活力,反之亦然。所以我们把玉米种子萌发期和苗期作为一个时期来研究,在这一阶段内,对玉米种子萌发和苗期生长条件容易控制,可多次重复,也具有时间短、容量大等优点,能反映玉米品种的抗旱性差异,进而评价不同玉米品种苗期的抗旱性也是比较科学的。本研究选用2个对干旱敏感性不同的玉米品种,通过精确控水法进行干旱胁迫处理,观察测定萌发期的农艺性状及苗期的生理指标,分析这2个玉米品种在种子萌发期及苗期对干旱胁迫的适应性,为玉米抗旱生产及品种选育提供参考。
1 材料和方法
1.1 试验材料
购自河南省农科院秋乐种子公司的登海605(DH)和伟科702(WK)2个玉米品种种子作为供试材料。
1.2 试验方法
试验于2018年7~10月在郑州师范学院温室进行。试验土壤取自河南农业大学科教园区试验田 0~30 cm 耕层土,土壤田间最大持水量为22.67%,基础肥力测定含有全氮 0.72 g.kg-1,有效磷 9.8 mg.kg-1,速效钾 124 mg.kg-1,有机质 9.59g.kg-1,碱解氮 17 mg.kg-1。萌芽期育苗钵选用底部有孔隙的塑料钵盘,每盘 50 穴,每穴体积为 34 cm3,覆膜选用透光性强的保鲜膜,通过精确称量育苗钵差量控水法计算得出土壤实际含水量[15]。对照组土壤相对含水量为田间最大持水量的75%,萌芽期处理组保持土壤含水量为田间最大持水量的 45%,为中度干旱胁迫 (依据《中华人民共和国水利行业标准——农业旱情等级划分标准》划分干旱胁迫梯度)。每个品种2组,每组3个生物学重复。供试材料于光照培养箱(上海智诚)内培养1周(日照时间为15 h,温度28℃,光照度6 000 lx;黑暗9 h,温度25℃),第8 d开始取样。
苗期先将玉米种子25℃黑暗处理3 d,催芽后移栽于草炭土壤中使其长至三叶一心期(期间正常浇水管理),生长17 d 后将长势一致的幼苗小心取出并将根冲洗干净分为两组,一组置于清水中培养作为对照(CK),一组置于20%PEG-600溶液中为干旱处理。24 h后测定光合及荧光参数,并取第3片(自下而上)迅速用液氮速冻,储存于-80℃冰箱中保存备用。
1.3 测定项目与方法
1.3.1 萌芽期生长量参数的测定 萌芽期生长指标测定及计算参照崔静宇等[15]的方法,连续一周期8 d,每天记录发芽粒数,第8 d开始取样,每个处理中挑选长势均匀一致的植株幼苗各6棵,将根小心从土壤中取出并冲洗干净,滤纸吸干表面水分后测量胚根主根长和株高。将上述取样植株分成地上、地下两部分置于105℃干燥烘箱中杀青 5 min,然后70℃恒温烘 12 h至恒重,用电子天平称量干质量。
1.3.2 苗期生理生化指标测定 利用便携式光合仪(LI-6400XT,USA)于上午10∶00选取相同位置(自植株底部第3片)完全展开叶测定各处理玉米幼苗的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2(Ci)浓度。用便携式可调荧光仪(PAM-2500,Heinz Walz GmbH,Germany)测定叶绿素荧光参数,包括最小荧光(Fo),最大荧光(Fm),光系统II(PSII)最大光化学效率(Fv/Fm)和电子传递速率(ETR)。化学比色法测定过氧化氢(H2O2)含量;硫代巴比妥酸法测定丙二醛(MDA)含量;质膜透性用电导率仪(DDSJ-308A, Shanghai, China)测定;氮蓝四唑法测定SOD活性;愈创木酚法测定POD活性;紫外分光光度计法测定CAT和APX的活性,及还原型谷胱甘肽(GSH)的含量;磺基水杨酸法测定游离脯氨酸含量;蒽酮比色法测定可溶性糖含量;考马斯亮蓝法测定可溶性蛋白含量;双吡啶法测定还原型抗坏血酸(AsA)的含量。
1.4 数据处理
利用Excel 2010进行原始数据整理;SPSS 20进行one way-ANOVA 分析,LSD法进行差异显著性检验。
2 结果分析
2.1 干旱胁迫对种子萌发各项指标的影响
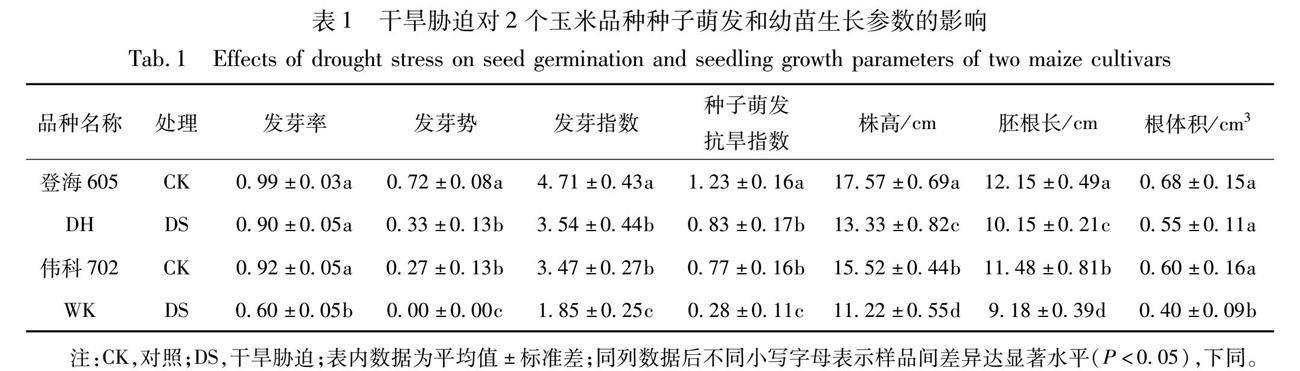
由表1中2个玉米品种种子发芽指标可以看出,中度干旱条件严重抑制玉米种子的发芽率、发芽势、发芽指数、萌发抗旱指数以及株高、胚根长和根体积,且存在品种间差异。与对照相比,干旱胁迫条件下DH玉米种子发芽率、发芽指数、萌发抗旱指数、株高、胚根长和根体积分别降低了9.09%,25.69%,32.65%,24.13%,16.46%,19.47%,而WK分别下降了34.78%,75.40%,63.29%,27.70%,20.03%,33.33%,其下降幅度明显高于DH;干旱胁迫条件下DH玉米种子相对发芽势为46.44%,而WK玉米种子相对发芽势为0%。由此得出,中度干旱胁迫抑制了玉米种子的发芽速率,且对2个玉米品种种子的发芽势较发芽率影响明显。本试验结果说明DH种子活力较高,在干旱条件下有较高的发芽率和一定的发芽优势,在低水分条件下发芽速率较快,对干旱胁迫的适应能力较强,比WK玉米品种在一定水分胁迫下较耐旱。此外,种子萌发抗旱指数现已被用于杂交种抗旱能力的鉴定[16]。从表1还可以发现,本试验中具有较高种子萌发抗旱指数的DH玉米种子发芽势受中度干旱胁迫影响较小,最终发芽率基本上没有显著地降低;而种子萌发抗旱指数较低的WK玉米品种则受中度干旱胁迫影响较大,其发芽势和发芽率都显著下降,说明该品种对中度干旱胁迫很敏感,该胁迫条件对它的种子萌发和幼苗生长产生了严重的抑制作用。
由表2可以看出,与对照相比,中度干旱胁迫使2个玉米品种在萌芽期的生物量积累明显受到抑制,且品种间差异显著。干旱条件下DH的地上干重、地下干重、总干重和贮藏物质转运率比CK分别降低了20.00%,31.91%,14.95%,23.24%,而WK比CK 分别降低了37.21%,38.71%,32.31%,33.33%,其减少程度明显高于DH各指标下降幅度。说明水分不足对种子萌发过程中的分解代谢和合成代谢活动产生了抑制作用,导致贮藏物质的无效消耗增多,利用转运效率降低,幼苗地上、地下干物质积累量也就相应减少。进一步说明中度干旱胁迫条件下WK种子在萌发过程中的分解及合成代谢生理活动受到了较严重的抑制,造成贮藏物质无效消耗较DH多,其地上、地下及总干重积累量较DH都低。另外,在相同处理条件下,2 個玉米品种的根冠比均有不同程度的升高,较耐旱品种DH幼苗的根冠比高于干旱敏感品种WK的根冠比。中度干旱胁迫下DH的根冠比比CK 增加了14.00%,WK比CK 增加了27.10%,更进一步说明WK地上部分干物质的积累较DH受干旱影响严重,导致根冠比升高幅度较大。
2.2 干旱胁迫对苗期叶绿素荧光和光合参数的影响
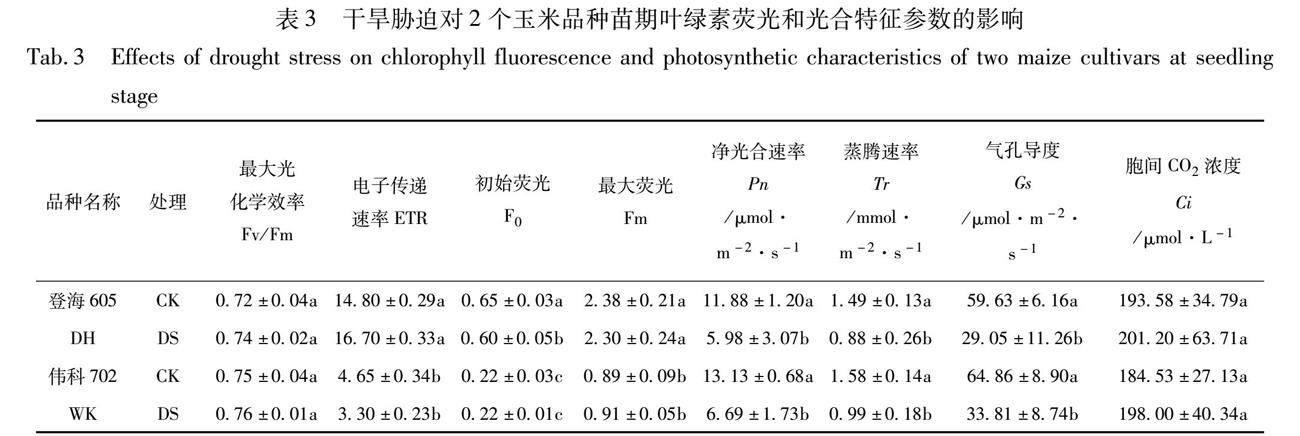
叶绿素荧光是灵敏快速、无伤害地研究植物光合作用与环境关系的内在探针,其参数可用来描述植物光合作用和光合生理状况。20%PEG-600溶液处理24 h后,2个玉米品种叶片叶绿素荧光参数Fv/Fm和Fm与各自的CK相比差异不显著(表3),但ETR在2个玉米品种间有显著差别,DH的ETR 在干旱处理条件下升高,而WK的ETR却降低。说明对2个品种幼苗进行24 h的20%PEG600溶液进行短时间的干旱处理,没有损坏玉米叶片的PSII结构,胁迫条件移去后,2个玉米品种幼苗均能较快的恢复正常生长和发育。已知F0为初始荧光,表示PSII 全部开放时的荧光产量,F0升高意味着PSII 受到了伤害或不可逆失活。Fm为最大荧光,表示PSII 反应中心处于完全关闭时的荧光产量,可反映经过 PSII 的电子传递情况[17]。本试验中,DH叶片的F0在干旱处理后下降,且在相同处理条件下,DH叶片的ETR和Fm参数高于WK,说明在干旱胁迫下DH幼苗叶片较WK有较强的光合电子转化能力和电子传递速率,表现出较好的光合性能。
光合作用影响作物生物量的积累,干旱等逆境胁迫一般会使玉米光合作用能力下降,导致植株生长减慢,而不同品种玉米在受到干旱胁迫后光合作用被抑制的程度不完全相同。由表3可以看出,干旱胁迫导致2个玉米品种苗期叶片的Pn,Tr和Gs均显著下降,DH的Pn,Tr和Gs比CK分别降低了49.66%,40.94%,51.28%,而WK比CK分别降低了49.05%,38.61%,47.87%,2个玉米品种叶片的Pn,Tr和Gs变化趋势基本一致。但是,在干旱处理条件下,2个品种叶片的Ci和对照比均差异不显著,稍微上升,说明2个品种叶片光合速率下降是非气孔因素所导致的。这可能是由于干旱胁迫导致Rubisco酶活性降低,对CO2亲和力下降,RuBP羧化酶再生能力降低以及叶绿体活性下降,最终导致了叶片的光合效率降低。
大量的研究表明,干旱等逆境条件会造成植物组织细胞内H2O2等ROS积累,进而引起膜脂质过氧化、膜结构及透性改变[18]。MDA是细胞膜脂质过氧化的最终产物,其积累量高低可表示细胞膜脂质过氧化作用的程度[19]。质膜透性是衡量细胞内容物扩散的一项生理指标,其数值变化大小可指示细胞质膜受到逆境伤害的程度[20]。由表4 数据可知,干旱胁迫处理使2个玉米品种叶片中的H2O2和MDA含量及细胞膜透性均显著升高,这说明干旱胁迫导致玉米叶片细胞内产生了较多的ROS,造成细胞膜脂质过氧化程度加剧,损伤了细胞膜的完整性,使细胞膜透性增大。与对照相比,干旱胁迫下DH叶中H2O2和MDA含量及细胞膜透性分别升高了24.37%,15.15%,9.00%,WK叶中分别升高了35.96%,19.79%,10.57%,DH叶中三者升高幅度均低于WK,3项指标都表明DH叶片细胞膜受干旱胁迫伤害程度较小,耐旱性较强。
抗氧化酶类主要有SOD,POD,CAT,APX,其中SOD通过歧化反应将O2-转化为H2O2,POD,CAT和APX将H2O2转化为O2和H2O,它们在清除过量ROS,抵抗氧化损伤过程中发挥着重要作用。由表4可知,在干旱处理条件下,2 个玉米品种叶片中的 SOD,POD,CAT,APX活性均升高。与对照相比,干旱胁迫下DH叶中SOD,POD,CAT和APX活性分别增加了8.60%,60.77%,61.77%,8.66%,WK分别增加了9.01%,24.68%,59.38%,48.51%。说明干旱胁迫诱导了玉米叶片中抗氧化酶基因的表达,合成了更多的抗氧化酶蛋白,从而减轻了ROS对细胞内生物大分子及细胞膜的伤害。其中,POD活性升高幅度在DH中远高于WK,而APX活性升高幅度在DH中远低于WK。另外,4 种抗氧化酶中,干旱胁迫使 DH叶中POD 和CAT的活性升高幅度较大,WK叶中CAT 和APX活性升高幅度较大,表明不同玉米品种叶中清除H2O2的主要酶类是有差异的。
脯氨酸、可溶性糖和可溶性蛋白是作物体内重要的渗透调节物质,在干旱胁迫条件下可以降低细胞渗透势,保持细胞膨压,提高其抗逆适应性[21]。如表5所示,干旱胁迫显著增加了玉米叶片中脯氨酸、可溶性糖和可溶性蛋白含量。与各自的对照相比,干旱胁迫使DH叶中3项指标分别升高了23.31%,17.56%,57.72%,WK叶中分别升高了12.29%,27.74%,7.12%。DH较WK叶中积累了较多的脯氨酸和可溶性蛋白来降低细胞渗透势,提高了细胞组织的持水作用,对细胞膜和细胞内容物起到更好的保护作用。干旱等逆境下可溶性糖含量增加也是作物对干旱胁迫的一种适应性反应,在相同干旱条件下DH叶中可溶性糖含量增加幅度稍低于WK,这说明并不是所有的渗透调节物质在较抗旱的玉米品种叶中升高幅度都较干旱敏感的品种高,这是由遗传机制形成的。
还原型抗坏血酸(AsA)和还原型谷胱甘肽(GSH)是普遍存在于植物体内物质,是H2O2的清除剂,在清除H2O2的抗坏血酸AsA系统中,AsA要在GSH的作用下才能还原成抗坏血酸,而继续清除细胞内过剩的H2O2,减少膜脂质过氧化[21]。由表5可知,在干旱处理条件下,2 个玉米品种叶片中的AsA和GSH含量都上升。与CK相比,干旱胁迫下DH叶中AsA和GSH分别升高了63.58% 和39.73%,升高幅度均高于WK,分别升高了24.62% 和27.29%。说明DH具有更强的清除H2O2的能力,更好地减轻ROS对细胞内生物大分子及细胞膜的伤害。裴英杰等[22]研究发现在水分胁迫条件下,细胞内AsA浓度升高,抗旱性越强的品种细胞内AsA浓度越高。研究还发现GSH能诱导与植物抗毒素生物合成有关的防御基因的转录和表达,高浓度的GSH有利于蛋白质的合成[23],我们的试验结果和上述结论一致,细胞内可溶性蛋白含量在抗旱的DH叶中积累量升高幅度比干旱敏感的WK大,更好地说明DH较WK耐干旱。
3 结 语
不同玉米品种在萌芽期对中度干旱胁迫的响应具有明显的差异,抗旱性较强的玉米品种在中度干旱缺水条件下仍能保持着相对较高的发芽势和发芽率,贮藏物质转运率所受干旱胁迫被抑制下降程度也較小,干物质积累量相对较高,这主要是由于不同材料的遗传基础和特性所决定的。干旱胁迫条件下, 2个玉米品种的光合作用特性和多项生理指标都发生了明显变化,在相同的指标中变化幅度表现出较大差异,但某项指标表现较好的品种在其他指标上表现却不一定突出,表明玉米的抗旱性也是多因素综合作用的复杂生理过程。总体上,我们的研究结果表明DH玉米品种在萌芽期有较强的种子活力,较高的种子萌发抗旱指数;在苗期表现出较强的渗透调节能力,其抗旱性较强,可以在后续试验中进行抗旱育种及栽培研究。
参考文献:
[1] FAROOQ M, WAHID A, KOBAYASHI N, et al. Plant drought stress: Effects, mechanisms and management[J].Agronomy for Sustainable Development, 2009, 29(1): 185-212.
[2] FAGHANI E, GHARECHAHI J, KOMATSU S, et al. Comparative physiology and proteomic analysis of two wheat genotypes contrasting in drought tolerance[J].Journal of Proteomics, 2015,114: 1-15.
[3] RIBAUT J M, JIANG C, GONZALEZ-DE-LEON D, et al. Identification of quantitative trait loci under drought conditions in tropical maize. 2. Yield components and marker-assisted selection strategies [J]. Theoretical and Applied Genetics, 1997, 94(6/7): 887-896.
[4] BOYER J S. Plant productivity and environment[J]. Science, 1982, 218(4571):443-448.
[5] 胡兴波,曹敏建,王学智,等.不同玉米品种萌芽期及苗期抗旱性初步研究[J].玉米科学,2004,12(3): 66-67.
HU X B, CAO M J, WANG X Z, et al. Studies on drought resistance of the different hybrid in seedling emergence stage and in seedling stage[J].Journal of Maize Science, 2004, 12(3): 66-67.
[6] 张仁和,马国胜,卜令铎,等.不同基因型玉米品种抗旱性鉴定及综合评价[J].种子,2009,28(10):91-94.
ZHANG R H, MA G S, BU L D, et al. Appraisition and comprehensive evaluation of different genotype maize cultivars for drought resistance[J].Seed, 2009, 28(10): 91-94.
[7] 高杰,张仁和,王文斌,等.干旱胁迫对玉米苗期叶片光系统Ⅱ性能的影响[J].应用生态学报,2015,26(5):1391-1396.
GAO J, ZHANG R H, WANG W B, et al. Effects of drought stress on performance of photosystem Ⅱ in maize seedling stage[J].Chinese Journal of Applied Ecology, 2015, 26(5): 1391-1396.
[8] 高盼,劉玉涛,王宇先,等.半干旱地区不同玉米品种的籽粒灌浆及脱水特性研究[J].黑龙江农业科学,2017 (12): 5-7.
GAO P, LIU Y T, WANG Y X, et al. Analysis of grain filling, dehydration characteristics of different maize varieties in semi-arid area[J].Heilongjiang Agricultural Sciences, 2017, (12): 5-7.
[9] 李运朝,王元东,崔彦宏,等.玉米抗旱性鉴定研究进展[J].玉米科学,2004,12(1): 63-68.
LI Y C, WANG Y D, CUI Y H, et al. Approach of study on maize drought-resistance identification[J].Journal of Maize Science, 2004, 12(1): 63-68.
[10]裴玉贺,轩慧冬,宋希云.不同玉米品种萌芽期抗旱筛选[J].山东农业科学,2017,49(2): 30-35.
PEI Y H, XUAN H D, SONG X Y, et al. Drought resistance screening of different maize varieties at germination stage[J].Shandong Agricultural Sciences, 2017, 49(2): 30-35.
[11]刘雪艳.不同基因型玉米品种抗旱性评价[D].杨凌:西北农林科技大学,2017.
[12]XU J, YUAN Y B, XU Y B, et al. Identification of candidate genes for drought tolerance by whole-genome resequencing in maize[J]. BMC Plant Biology, 2014, 14(1):1-15.
[13]MAHESWARI M, TEKULA V L, YELLISETTY V, et al. Functional mechanisms of drought tolerance in maize through phenotyping and genotyping under well watered and water stressed conditions[J]. European Journal of Agronomy, 2016, 79(5):43-57.
[14]NUCCIO M L, WU J, MOWERS R, et al. Expression of trehalose-6-phosphate phosphatase in maize ears improves yield in well-watered and drought conditions[J]. Nature Biotechnology, 2015, 33(8): 862-869.
[15]崔静宇,关小康,杨明达,等.基于主成分分析的玉米萌发期抗旱性综合评定[J].玉米科学,2019, 27(5): 62-72.
CUI J Y, GUAN X K, YANG M D, et al. Integrative evaluation of maize drought tolerance in germination period by PCA method[J].Journal of Maize Science, 2019, 27(5): 62-72.
[16]徐田军,吕天放,赵久然,等.玉米萌发幼苗期的抗旱性鉴定评价[J].中国种业,2017(4): 42-46.
XU T J, LYU T F, ZHAO J R, et al. Evaluation on drought resistance of maize at germination and seedling stage[J].Chinese Seed Industry, 2017(4): 42-46.
[17]吕丹丹,裴孝伯.外源亚精胺、壳聚糖对盐胁迫下黄瓜幼苗叶绿素荧光参数的影响[J].农业科学,2014(4):28-37.
LYU D D, PEI X B. Effect of exogenous spermidine (Spd), chitosan (CTS) on chlorophyll fluorescence parameters of cucumber seedling under salt stress[J].Agricultural Science, 2014, 4(2): 28-37.
[18]郭竞选,彭 雷,樊若楠,等.玉米‘郑58与‘PH4CV苗期干旱耐性比较分析[J].中国农学通报,2017 33(35): 18-24.
GUO J X, PENG L, FAN R N, et al. Comparative analysis of drought tolerance at seedling stage between maize Zheng58 and PH4CV[J].Chinese Agricultural Science Bulletin, 2017, 33(35):18-24.
[19]范晶,黄明远,徐雁霞.盐胁迫对番茄种子萌发及叶片中丙二醛含量的影响[J].北方园艺,2011,25(10):27-29.
FAN J, HUANG M Y, XU Y X. The effects of salt stress on tomato seeds germination and malondidehyde content in the leaf of the cultivated tomato[J].Northern Horticulture, 2011, 25(10): 27-29.
[20]原立地.DCPTA增强玉米苗期耐冷性的生理机制调控效应[D].哈尔滨:东北农业大学,2013.
[21]李凤海.玉米抗旱性指标的筛选及其遗传特性研究[D].沈阳:沈阳农业大学,2011.
[22]裴英杰,郑家玲,庾红,等.用于玉米品种抗旱性鉴定的生理生化指标[J].华北农学报,1992,7(1):31-35.
PEI Y J, ZHENG J L, YU H, et al. The indexes of physiology and biochemistry used for appraisal and level determination of drought resistance in maize[J].Acta Agriculturae Boreali-Siniea, 1992, 7(1):31-35.
[23]陈少裕.植物谷胱甘肽的生理作用及其意义[J].植物生理学通讯,1993,29(3):210-214.
CHEN S Y. Physiological roles of glutathione in plant and its significances[J].Plant Physiology Communications, 1993, 29(3): 210-214.
(编 辑 李 波,邵 煜)
猜你喜欢
江苏农业科学(2016年8期)2017-02-15 19:38:57
江苏农业科学(2016年8期)2017-02-15 19:26:11
山东农业科学(2016年12期)2017-01-21 14:41:54
山东农业科学(2016年12期)2017-01-21 14:39:43
安徽农学通报(2016年24期)2017-01-12 21:06:57
安徽农学通报(2016年24期)2017-01-12 21:04:01
现代园艺(2016年7期)2017-01-09 14:06:28
山东农业科学(2016年11期)2016-12-17 21:16:16
吉林农业·下半月(2016年10期)2016-12-16 23:45:53
现代园艺(2016年2期)2016-03-15 16:05:02
