
不同小麦品种对晚霜冻的响应及抗霜性评价
2020-04-29 02:35席吉龙席天元张建诚姚景珍席凯鹏李永山贾秀领
干旱地区农业研究 2020年1期
席吉龙,王 珂,杨 娜,席天元,张建诚,姚景珍,席凯鹏,李永山,贾秀领
(1.山西省农业科学院棉花研究所,山西 运城 044000;2.河北省农林科学院粮油作物研究所,河北 石家庄 050035)
霜冻害是小麦生产上的主要气象灾害之一。小麦完成春化后生长发育加快,随着幼穗分化期的推进,对低温的敏感性增加,幼穗的抗霜力总体呈下降的趋势[1-3]。小麦晚霜冻主要发生在春季3月下旬到4月上中旬,此时幼穗迅速发育,是影响穗粒数和产量的关键时期。发生晚霜冻致使叶片、穗下茎、幼穗不同程度受害,幼穗部分死亡或全穗枯死 ,造成不同程度的减产[4-5]。小麦霜冻年际发生频率为30%~40%[6]。在气候变暖背景下,一方面小麦种植区域向北和向高海拔地区推移扩大,品种选择偏春性化,小麦拔节抽穗期缩短,抽穗提早,抗寒力降低。另一方面异常低温气候趋多趋强[7-8],致使晚霜冻的成灾面积上升、危害程度加重[9],对小麦生产构成严重威胁。小麦晚霜冻害的发生与气候、土壤、植株营养和水分、栽培方式等有关,但品种抗寒性是关键性内在因素[10]。选育抗霜性强的品种是减轻霜冻害的重要途径,因此鉴定品种的抗霜性对小麦生产具有重要意义。
小麦抗霜性是小麦受到低于0℃及以下的低温胁迫后,通过一系列复杂的生理生化反应及形态结构的变化,对低温环境进行响应[11-12]。研究表明,植物的抗寒性与叶绿素含量[13]、光合作用[14-15]、叶绿素荧光[15-16]、细胞膜的通透性[17]、膜脂过氧化程度[15]、抗氧化酶活性[19]、丙二醛含量[20]、可溶性蛋白含量[20]密切相关。前人常用这些生理生化指标鉴定品种抗寒性,并采用多种指标来综合评价作物的抗寒性。隶属函数法是综合评价作物抗逆性的常用方法[20],能比较准确地反映出多个品种间的抗寒性差异。目前,在模拟低温盆栽环境下,通过生理生化指标对小麦的抗寒性评价研究较多[21-22],少有自然低温大田环境下,用生理生化指标结合幼穗形态及产量进行的抗寒性研究。本研究以黄淮海麦区北片三个不同类型、具有典型特点的16个主栽新品种为材料,在拔节期自然低温霜冻环境下,分析叶片生理生化变化和植株形态变化。以霜冻前后各性状的相对值作为抗霜性评价指标,通过隶属函数法和聚类分析法对参试品种抗霜性进行综合鉴定与评价,筛选出抗霜性品种及抗霜性鉴定指标,为气候变暖背景下小麦品种选育与抗霜减灾提供理论支持。
1 材料与方法
1.1 试验地概况
试验在山西省农业科学院棉花研究所牛家凹农场进行,试验所在地属黄河流域西北部小麦区,试验地前茬为小麦,土壤为壤质土,肥力均匀,播前0~20 cm耕层土壤有机质12.36 g·kg-1,全氮0.74 g·kg-1,有效磷9.5 mg·kg-1,速效钾174.4 mg·kg-1。播前基施尿素293.5 kg·hm-2,重过磷酸钙326.0 kg·hm-2,返青期追施尿素195.5 kg·hm-2。
1.2 试验材料
供试小麦品种及其选育单位如表1所示。
1.3 试验设计
以16个小麦品种为16个处理,每个品种以霜冻前为对照重复3次,随机区组排列,小区面积15 m2。2017年10月8日播种,行距25 cm,各处理基本苗控制在255万株·hm-2,依据千粒重、发芽率及田间出苗率计算播量,水肥等田间管理一致,于返青期和开花期各灌水60 mm。2018年4月7日凌晨试验地出现自然低温,温度为-2.6℃~-1.1℃,持续时间5 h,4月6日20∶00至4月7日10∶00气温变化见图1。
1.4 测定内容与方法
1.4.1 基本调查 在小麦主要生育期调查群体结构及个体发育情况。在春季霜冻后进行植株形态、幼穗冻伤率等调查。成熟期调查产量和产量结构。

表1 供试小麦品种及其选育单位

图1 晚霜冻期间的气温变化Fig.1 Temperature changes during frost
1.4.2 生理生化指标测定 在晚霜冻来临前3 d和晚霜冻后3 d对各品种分别进行以下测定。叶绿素含量用日本SPAD-502叶绿素仪测定,每小区选5株测定上部第1片完全展开叶,每个叶片测定一次。叶绿素荧光参数用美国OS-30P叶绿素荧光仪测定,每小区选5株测定上部第1片完全展开叶,于晴天上午9∶00—12∶00测量,测量前将叶片预先暗适应30 min,测PSⅡ原初光能转化效率(Fv/Fm),取5片叶的平均值作为测定值。光合作用采用英国生产的LCPRO便携式光合测定仪[23],每小区随机选取上部第1片完全展开叶3片,在叶片相同部位测定,每个叶片测定3 min 左右,测得小麦光合速率(Pn)等参数,取3片叶的平均值分析使用。过氧化物酶(POD)活性采用愈创木酚比色法[24],过氧化氢酶(CAT)活性测定采用紫外吸收法[21],超氧化物歧化酶(SOD)活性采用NBT比色法[24],丙二醛(MDA)含量采用TBA比色法[21],相对电导率采用电导率仪法[24]。
1.5 数据分析
1.5.1 性状相对值 以公式(1)计算性状抗冻系数相对值(xj),作为评价品种抗霜性的指标:
xj=低温处理下某指标测定值/对照测定值×100%
(1)
1.5.2 相对性状的隶属度值 以公式(2)和公式(3)计算各品种相对性状的隶属度值[25]。其中,与抗寒性呈正相关的各指标(叶绿素、叶绿素荧光、光合速率、SOD活性、POD活性、CTA活性)隶属度值依据公式(2)计算,与抗寒性呈负相关的各指标(MDA含量和相对电导率)隶属度值依据公式(3)计算。每一品种相对性状隶属度值的平均数即为该品种的平均隶属度值:
U(xij)=(xij-xjmin)/(xjmax-xjmin)
(2)
U(xij)=1-(xij-xjmin)/(xjmax-xjmin)
(3)
式中,U(xij)为i品种j指标的隶属函数值,xij为i品种j指标的相对值,xjmin和xjmax为各品种j指标相对值的最小值和最大值。
1.5.3 隶属度平均值评价法 以多个性状的隶属度平均值作为评价指标,平均隶属度值高,该品种的抗寒性强;反之,该品种的抗寒性弱。
1.6 统计分析
采用Microsoft Excel 2007软件对数据进行处理,采用DPS 7.05数据分析软件进行方差分析(用新复极差法进行多重比较)、聚类分析。
2 结果与分析
2.1 晚霜冻对小麦叶片生理指标的影响
2.1.1 相对电导率 在霜冻来临前的正常生长条件下,参试16个小麦品种叶片相对电导率平均为33.5%(图2),以晋麦84号叶片相对电导率表现最高,为40.6%,以冀麦325最低,为25.5%。霜冻发生后,叶片细胞膜受损电解质渗出增加,相对电导率平均高出对照32.8%,品种间差异极显著(P<0.01),不同品种小麦叶片相对电导率变动幅度比霜冻前增加。在供试的16个品种中,以霜冻前为对照,周麦18叶片相对电导率增幅最高,为47.7%,而济麦22增幅最低,仅为16.6%。
2.1.2 叶绿素相对含量(SPAD值) 叶绿素在光合作用中起吸收光能的作用,其含量直接影响到光合作用。霜冻前对照组叶绿素相对含量SPAD值平均为57.56,以西农529的值最高,为60.68,临Y8012叶绿素含量最低,为52.57(图3)。霜冻后叶绿素含量平均为54.18,比对照低5.9%,品种间差异极显著(P<0.01)。与霜冻前相比,西农529叶片叶绿素含量降低幅度最大,降低了15.6 %,而烟农1212仅降低了0.6 %。表明晚霜冻使叶绿素含量下降,叶绿素含量的下降主要是由于叶绿素合成受到抑制。

图2 拔节期霜冻害后小麦叶片相对电导率Fig.2 Relative electrical conductivity in wheat leaves after spring frost injury

图3 拔节期霜冻害后小麦叶片叶绿素相对含量Fig.3 Relative chlorophyll content in wheat leaves after spring frost injury
2.1.3 叶绿素荧光参数(Fv/Fm) 由图4看,霜冻前对照组叶绿素荧光参数(Fv/Fm)平均为0.821,以周麦18的值最高,为0.836,以冀麦325的最低,为0.812。霜冻后小麦叶片叶绿素素荧光参数(Fv/Fm)平均值为0.813,平均降低1.0%,品种间差异显著(P<0.05),降幅最大的西农529,叶片叶绿素含量降低了2.9%,而烟农1212仅降低了0.3%。
2.1.4 光合速率 光合速率是反映植物光合作用强弱的重要指标。由图5看,在霜冻来临前的正常生长条件下,参试的16个小麦品种叶片光合速率平均为14.63 μmol·m-2·s-1(图5),以山农30叶片光合速率最高,为16.09 μmol·m-2·s-1,以冀麦325最低,为11.94 μmol·m-2·s-1。霜冻后叶片光合速率平均降低34.4%,不同品种光合速率变动幅度比霜冻前增加,品种间差异显著(P<0.05)。在供试的16个品种中,各品种霜冻后与霜冻前(对照)相比,科农2009叶片光合速率降幅最高,为44.0%,而山农28降幅最低,仅为26.4%。
2.2 晚霜冻对小麦保护酶及丙二醛的影响
2.2.1 POD活性 正常生长条件下,参试的16个小麦品种POD活性平均为50.5 U·g-1(图6),以西农585叶片POD活性最高,为61.12 U·g-1,以衡杂102最低,为44.4 U·g-1。霜冻后叶片POD活性平均为67.5 U·g-1,POD活性高出对照33.7%,品种间差异极显著(P<0.01)。与对照相比,在供试的16个品种中,济麦23叶片POD活性增幅最高,为61.9%,而西农585增幅最低,仅为15.7%。

图4 拔节期霜冻害后小麦叶片荧光参数Fig.4 Fluorescence parameters (Fv/Fm) of wheat leaves after spring frost injury
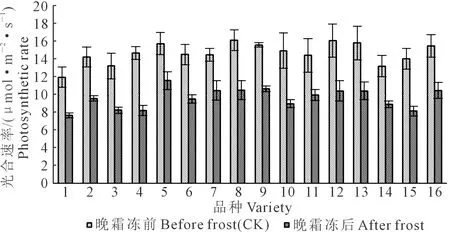
图5 拔节期霜冻害后小麦叶片光合速率Fig.5 Photosynthetic rate of wheat leaves after spring frost injury
2.2.2 CAT活性 正常生长条件下16个小麦品种CAT活性平均为2 735.4 U·g-1(图7),以山农30叶片CAT活性最高,为3 132.4 U·g-1,以晋麦84号最低,为2 486.6 U·g-1。霜冻后叶片CAT活性平均为2 966.1 U·g-1,CAT活性高于对照8.4%,品种间差异显著(P<0.05)。与对照相比,在供试的16个品种中,济麦23号叶片CAT活性增幅最高,为21.6%,而山农30却降低4.5%。
2.2.3 SOD活性 正常生长条件下16个小麦品种SOD活性平均为277.4 U·g-1(图8),以西农529叶片SOD活性最高,为354.8 U·g-1,以师栾02-1最低,为234.4U·g-1。霜冻后叶片SOD活性平均为406.9 U·g-1,SOD活性高于对照46.7%,品种处理间差异极显著(P<0.01)。与对照相比,在供试的16个品种中,济麦23叶片SOD活性增幅最高为83.8%,而科农2009仅提高了8.8%。参试小麦品种叶片SOD活性均有提高,说明小麦增强了对低温逆境的适应能力,且不同小麦品种对逆境的适应能力不同。
2.2.4 MDA含量 正常生长条件下16个小麦品种的MDA含量平均为20.52 nmol·g-1(图9),以晋麦84MDA含量最高,为33.25 nmol·g-1,以临Y8012最低,为16.76 nmol·g-1。霜冻处理后,MDA含量平均为24.79 nmol·g-1,比对照提高20.8%,品种间差异极显著(P<0.01)。与对照相比,在供试的16个品种中,小麦功能叶MDA含量变化不同,科农2009MDA含量增幅最高,为39.2%,而山农28提高了7.2%。抗霜性较强的小麦品种MDA积累少、膜质过氧化程度低,抗霜性较弱的小麦品种MDA积累多、膜质过氧化程度高。MDA含量的增加显示霜冻对小麦伤害程度较重。
2.3 晚霜冻对小麦产量构成因素及产量的影响
小麦在拔节中后期遇自然霜冻后,起初穗下节和幼穗出现水渍状水肿,冻害程度不易分辨,抽穗后穗部冻害症状逐渐明显,4月25日调查结果表明(表2),不同品种幼穗死伤率差异显著,幼穗死伤率在1.3%~41.8%。按幼穗死伤率高低分为3类:幼穗死伤率低的有济麦22、山农28、济麦23、中麦36、临Y8012、舜麦1718、烟农1212、山农30;幼穗死伤率高的有西农529、周麦18;其余幼穗死伤率属中间类。从表2可见,拔节期遇霜冻造成幼穗死伤,从而有效穗数和穗粒数减少,对不同小麦品种的产量和产量结构影响显著,产量4 328.1~8 222.3 kg·hm-2。说明不同品种的抗霜性存在差异,拔节期霜冻减产的主要原因是有效穗数和穗粒数的减少。

图8 拔节期霜冻害后小麦叶片SODFig.8 SOD activity in wheat leaves after spring frost injury
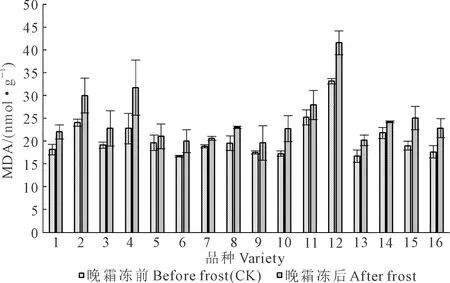
图9 拔节期霜冻害后小麦叶片MDAFig.9 MDA content in wheat leaves after spring frost injury

表2 晚霜冻对小麦产量和产量构成因素的影响
注:同列不同字母表示P<0.05水平下差异显著(P<0.05)
Note: Values followed by different letters in a column were significantly different treatments atP<0.05 level.
2.4 小麦拔节期霜冻生理生化指标的隶属度法评价
小麦抗冻性是一个复杂的综合性状,用单项指标评价都有片面性,需多指标综合评价。根据多项生理指标的测定值,用公式(1)求出各单项指标的抗冻系数。利用公式(2)、(3)进一步计算每一品种的平均隶属度值,根据各品种的隶属度平均值D(表3),评价品种抗冻能力的大小。对参试品种抗冻性进行强弱排序,其中山农28的D值最大,表明该品种抗冻性最强,其次为济麦22、济麦23、临Y8012、中麦36、舜麦1718麦等,西农529、周麦18、科农2009、西农585、师栾02-1、冀麦325的D值较小,表明其抗冻性差。
2.5 小麦拔节期抗晚霜性综合评价
采用最长距离法,以各品种8个性状的隶属度平均值、幼穗冻伤率、产量三个指标进行聚类分析,结果分为五类(见图10):I类:济麦22、临Y8012、山农28;II类:舜麦1718、济麦23、中麦36、烟农1212;III类:山农30、晋麦84号、衡杂102、师栾02-1、科农2009、周麦18;IV类:冀麦325、西农585;V类:西农529。从第I类品种到第V类品种,冬性越来越弱,春性越来越强,抗晚霜冻能力越来越弱。

表3 晚霜冻后小麦品种综合隶属度值

图10 参试小麦品种抗晚霜冻聚类分析Fig.10 Cluster tree chart of wheat varieties on frost resistance
3 讨 论
植物为适应低温胁迫环境通过细胞一系列生理生化变化而进行自我调节[11]。细胞膜是细胞与环境发生物质交换的主要通道,当质膜受低温逆境伤害时,膜的通透性发生变化,常以叶片电导率作为鉴定细胞膜伤害程度和所测材料抗逆性的大小[25]。本研究表明,拔节期小麦受到霜冻胁迫后叶片相对电导率较对照显著增加。因此,霜冻使小麦叶片膜透性增加并且膜受损程度加大。
陈思思[14]、王瑞霞[15]等研究表明,低温胁迫下叶片绿素含量、叶绿素荧光参数(Fv/Fm)、光合速率发生变化。本研究表明,霜冻胁迫后叶片叶绿素含量下降,直接影响叶绿素荧光参数和光合速率显著下降。
低温胁迫打破了细胞内自由基的产生和清除平衡状态,自由基过多,引发膜脂过氧化作用[26],产生有害的过氧化产物MDA。随温度降低和胁迫时间的延长膜脂过氧化作用加重[27],然而,植物体中还存在着SOD、POD等抗氧化酶防御系统,以清除活性氧、降低逆境损伤[28]。本研究表明,小麦拔节期霜冻使小麦叶片SOD活性、POD活性、CAT活性和MDA含量均高于对照。SOD活性、CAT活性和POD活性的增加可以清除自由基减轻低温对生物膜的伤害。但是MDA含量的增加显示膜脂过氧化作用加重,即霜冻对小麦伤害程度较重。
倒春寒对小麦产量及产量构成的影响已有相关研究[15,29-30]。在高于突变温度区间的晚霜冻胁迫下,穗粒数的下降是产量损失的主要原因,在低于突变温度区间的晚霜冻胁迫下,不仅穗粒数下降,而且部分植株死亡,使穗数也显著降低[29-30]。王瑞霞等[15]认为拔节期和孕穗期低温胁迫可导致小麦茎、幼穗受冻,穗数和穗粒数显著减少,小麦产量均显著下降。胡新等[31]研究认为每百穗中各级冻害穗数、穗粒数和实际籽粒重均显著低于未受冻穗,因此导致小麦减产。本研究表明,参试小麦各品种的幼穗均受到不同程度的伤害,幼穗冻伤率1.3%~41.8%,说明不同小麦品种抗霜冻特性存在差异。受冻小麦幼穗表现为不抽穗、空心穗、缺粒穗,导致穗数、穗粒数和籽粒产量降低。不同小麦品种间产量和产量构成差异显著。本研究还发现,主茎和大分蘖的幼穗冻死后,晚生蘖(再生分蘖)快速生长可以成穗和结实,但16个品种的晚生蘖穗粒数平均降低45.1 %,千粒重降低36.1 %,单穗重平均降低了65.1 %。晚生穗理论上可弥补霜冻灾害带来的部分产量损失,实际上晚生穗比正常穗成熟期推迟5~7 d,无法实施机械统一收获,混收后造成数量多而品质降低的情况。
在对多个抗霜性指标进行更加准确的综合评价时,目前多用主成分分析法[18]、隶属函数法[25]、聚类分析[32]等方法,其中隶属函数法在作物抗寒性评价方面应用较多。本试验应用多指标相对性状值,计算出各品种的平均隶属度值,进行了综合评价。并通过平均隶属度值、幼穗冻伤率、产量聚类,将供试材料聚为5个类群,通过隶属度平均值(D值)、幼穗冻伤率和产量聚类,将16个小麦品种聚为5类,I类:济麦22、临Y8012、山农28;II类:舜麦1718、济麦23、中麦36、烟农1212;III类:山农30、晋麦84号、衡杂102、师栾02-1、科农2009、周麦18;IV类:冀麦325、西农585;V类:西农529。从第I类品种到第V类品种,冬性越来越弱,春性越来越强,抗晚霜冻能力越来越弱。
综合评价结果客观反映了品种的抗霜性。D值的排序与聚类分析有较高的吻合度。单一运用D值排序虽可得到鉴定材料的抗寒性综合评价,却无法得到鉴定材料之间的遗传关系,也不容易对抗寒性进行分类或分级;仅用聚类分析虽然能较好地反映鉴定材料在抗寒性方面的遗传关系,却易对鉴定材料抗寒敏感性结果造成偏差。只有将两者有机地结合使用,才能较为全面地对供试材料进行抗寒级别划分和抗寒性评价。因此,在开展作物抗寒性评价时,建议将隶属函数法和聚类分析方法结合使用。
培育优良抗低温品种是应对小麦晚霜冻的一条经济有效的途径,田间品种鉴定结果为小麦抗逆减灾和育种提供了重要依据。在田间鉴定的基础上,结合位点基因型的分子标记鉴定, 深入研究小麦抗寒性基因,揭示基因在小麦抗寒性中的作用及机制。开展田间鉴定、常规育种、基因工程、细胞工程育种等多个学科相结合的系统研究,培育出适应未来气候变化的优良品种。
4 结 论
在小麦拔节期遇自然霜冻(-2.6℃~-1.1℃)突袭5 h,16个不同小麦品种拔节期功能叶叶绿素含量、叶绿素荧光参数(Fv/Fm)、光合速率下降,叶片SOD活性、POD活性和CAT活性、相对电导率均不同程度地上升。通过隶属度平均值(D值)、幼穗冻伤率和产量聚类,将16个小麦品种聚为5类,I类:济麦22、临Y8012、山农28;II类:舜麦1718、济麦23、中麦36、烟农1212;III类:山农30、晋麦84号、衡杂102、师栾02-1、科农2009、周麦18;IV类:冀麦325、西农585;V类:西农529。从第I类品种到第V类品种,冬性越来越弱,春性越来越强,抗晚霜冻能力越来越弱。隶属函数法和聚类分析方法结合使用是进行小麦抗霜性综合评价的有效方法。
猜你喜欢
华北农学报(2022年4期)2022-09-14
热带作物学报(2022年5期)2022-06-01
东北农业大学学报(2020年12期)2021-01-15
今日农业(2020年20期)2020-12-15
今日农业(2020年20期)2020-12-15
山西农业科学(2020年10期)2020-10-16
浙江农业学报(2020年4期)2020-04-28
江苏农业科学(2019年10期)2019-07-08
江苏农业科学(2016年11期)2017-03-21
