
三峡库区消落带典型植物光合固碳能力及影响因素
2020-04-27 08:06冯晶红肖衡林吴耕华王可健
水土保持研究 2020年1期
冯晶红, 刘 瑛, 肖衡林, 刘 煜, 吴耕华, 王可健
(湖北工业大学 土木建筑与环境学院, 河湖生态修复及藻类利用湖北省重点实验室, 武汉 430068)
水库消落带又称涨落带或涨落区,指的是水库季节性水位涨落而使周边被淹没土地周期性地出露于水面的一段特殊区域[1-2],是水生生态系统和陆生生态系统交替控制的过渡地带,是一类特殊的湿地生态系统[3],在控制陆地生态系统碳循环中扮演着重要的角色,即湿地可以从大气中吸收二氧化碳并固定成为碳库,同时又向大气中排放CH4,成为碳源[4]。据研究表明,三峡水库消落带出露面积达437~446 km2,如此大面积的消落带植被吸收了大量CO2,其积累的生物物质是宝贵的资源[3-5]。因此,需要重视三峡水库消落带湿地植被作为碳库在储存有机碳中的重要作用。
植物作为碳库在储存有机碳中发挥着重要作用,主要体现在植物具有固碳释氧的重要功能,即通过光合作用吸收二氧化碳和释放氧气[6],对调节区域碳平衡起着重要的作用[7]。目前对于湿地生态系统植物固碳能力方面,国内外学者已经开展了广泛而深入的研究。Crill[8]等通过对北方泥炭地湿地植物的研究,得出其固碳能力约为0.31 kg/(m2·a);Aselmann[9]等通过研究认为全球湿地植物的平均固碳能力为0.05~1.35 kg/(m2·a);我国许多学者也对不同类型湿地生态系统的植被的储碳和固碳能力进行了研究[10-11],而鲜有对于消落带这类特殊湿地植被的储碳和固碳能力的研究。基于以上考虑,本研究拟从不同植物光合特性和固碳能力的角度出发,选取了三峡库区消落带地区15种典型适生植物,对其光合固碳能力进行分析比较,定量化探究三峡库区消落带优势植物的光合固碳能力,同时对光合固碳能力的关键性指标净光合速率的内外影响因子进行相关分析,为三峡库区消落带生态恢复植物筛选和高效碳汇植物配置提供科学依据,同时为其他大型水库应对碳循环问题提供参考。
1 材料与方法
1.1 试验区域概况
试验样地位于重庆三峡库区开州区至万州区段人工恢复消落带。位于长江上游下段,全区地处中纬度,属湿润亚热带季风气候,多年平均气温15~18℃,多年平均降雨量为1 150.26 mm,表现出明显的年、季变化。土壤类型主要有水稻土、黄壤、石灰土、紫色土等。植被是一年生草本为主的草丛植被类型,灌丛植被分布较少,且仅有少量乔木幼树或萌生枝条,主要有狗牙根(Cynodondactylon)、辣蓼(Polygonumhydropiper)、苍耳(Xanthiumsibiricum)、鬼针草(Bidenspilosa)、芦竹(Arundodonax)、大狼把草(Bidensfrondosa)、黄荆(Vitexnegundo)、桑(Morusalba)、枫杨(Pterocaryastenoptera)、刺槐(Robiniapseudoacacia)、垂柳(Salixbabylonica)、毛白杨(Populustomentosa)等。
1.2 试验材料
试验选取从开州区渠口镇至万州区石宝寨人工恢复消落带区段作为试验样地。该区每年10月至次年1月高水位运行(水位高于海拔170 m),于6月至8月低水位运行(水位低于海拔150 m)。为了减少地形地貌、水位波动及气候变化等因素对植被光合作用的影响,在植物正处于夏季旺盛生长季(7月下旬)开展调查研究。根据植物群落组成特征,选取了近自然状态的15种典型消落带适生植物,分别为中山杉(Taxodium‘Zhongshanshan’)、竹柳(Salixsp)、柑橘(Citrusreticulata)、苎麻(Boehmerianivea)、牡荆(Vitexnegundovar.cannabifolia)、桑(Morusalba)、地桃花(Urenalobata)、苍耳(Xanthiumsibiricum)、风车草(Cyperusalternifolius)、芦苇(Phragmitesaustralias)、白芒(Miscanthussinensis)、大狼把草(Bidensfrondosa)、鬼针草(Bidenspilosa)、合萌(Aeschynomeneindica)、辣蓼(Polygonumhydropiper),隶属于12科,分为3种生活型:乔木3种、灌木4种、草本8种。
2 试验方法
2.1 植物光合速率和叶面积指数的测定
植物光合速率测定采用SY-1020便携式光合仪。于2017年7月20日至7月30日选择光照充足、无风或微风的晴天,在自然光照条件下,从8:00—18:00每隔2 h测定1次,并同步测定空气气温(Ta)、叶片温度(TL)、叶室湿度(RH)、光合有效辐射(PAR)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)和WUE(水分利用率)等参数。测定时每种植物选择具有代表性的3株植株,每次每株植物选择向阳的大小相近、长势较好的功能叶4~6片,在不离体的情况下每片叶记录5个瞬时光合速率值,然后取其平均值,每种植物取3株样木的平均值,各个时间段内测定树种的顺序保持不变。
利用S-120型便携式叶面积测量仪对所选植物的叶面积进行测定。每种植物选取3株具有代表性的样株,每株植物测量6片单叶,并取其均值作为该种植物的单叶叶面积,同时采用标准枝法调查整株叶量,进而获得整个调查植株的叶片总面积(Y)[12]。植物的冠幅大小(S)用皮尺测定,其中乔木和灌木选取4株样株测定其冠幅大小,取平均值,草本植被覆盖面积以1×1 m2样方计数。每种植物单株叶面积指数(leaf area index,简称LAI)计算公式[6]为:
LAI=Y/S
(1)
2.2 日净同化总量和固碳能力的计算及分析方法
利用简单积分法计算测试植物在测定当日的净同化量[13-14],计算公式为:
(2)
式中:P为测定日单位叶面积的净同化总量[mmol/(m2·d)];pi为初测点的瞬时光合作用速率;pi+1为i+1测点的瞬时光合作用速率[umol/(m2·s)];ti为初测点的瞬时时间;ti+1为i+1测点的时间(h);j为测试次数。
一般植物晚上的暗呼吸消耗量按照白天同化量的20%计算[15],则单位叶面积日净固定CO2量按照以下公式计算:
WCO2=p×(1-0.2)×44/100
(3)
式中:44为CO2的摩尔质量(g/mol);WCO2为测定日单位叶面积的净固定CO2的质量[g/(m2·d)]。
测试植物在测定当日单位土地面积上每天吸收CO2按照以下公式计算:
QCO2=WCO2×LAI
(4)
式中:QCO2为测定日单位土地面积的净固定CO2的质量[g/(m2·d)]。
2.3 叶片氮磷含量的测定
叶片样品自野外带回实验室后进行杀青,烘干处理,然后对叶片营养元素进行测定。叶片全氮含量利用凯氏定氮仪(FOSS 2200,瑞典);植物叶片全磷含量采用H2SO4—H2O2消煮—钼锑抗比色法测定的[16]。其中TN,TP定义为单位重量叶片的全氮、全磷含量。
2.4 数据分析
利用SPSS 22.0数理统计软件对数据进行相关分析和方差分析。不同生活型植物叶片光合特性与影响因素及叶片养分含量之间的关系采用Pearson相关分析法。
3 结果与分析
3.1 净光合速率与主要环境生理因素相关性分析
见表1,不同植物净光合速率对自然环境因子的响应不相同。15种植物的净光合速率与光合有效辐射(PAR)、蒸腾速率(Tr)及气孔导度(Gs)等呈正相关,且大部分测试植物的净光合速率与光合有效辐射(PAR)、蒸腾速率(Tr)及气孔导度(Gs)呈显著(p<0.05)或(p<0.01)极显著的正相关。而测试植物的净光合速率与胞间CO2浓度(Ci)呈负相关,且部分植物呈显著(p<0.05)或(p<0.01)极显著的负相关。
测试的大部分植物的净光合速率与气温(Ta)、叶片温度(TL),叶室湿度(RH)以及水分利用(WUE)成正相关,并且部分植物的净光合速率基本上与气温(Ta)、叶片温度(TL),叶室湿度(RH)以及水分利用(WUE)呈显著(p<0.05)或(p<0.01)极显著的正相关;而苎麻、白芒、大狼把草及鬼针草的净光合速率则与气温(Ta)、叶片温度(TL),叶室湿度(RH)以及水分利用(WUE)呈负相关,其中白芒及大狼把草净光合速率则与气温(Ta)、叶片温度(TL)呈显著(p<0.05)或(p<0.01)极显著的负相关。
3.2 植物叶片的日平均光合速率与叶片氮、磷含量的相关性分析
3.2.1 不同生活型植物日平均光合速率与叶片N,P含量分析 氮磷对植物光合作用及碳分配有着显著的影响[17],故而影响植物光合固碳能力。由表2看出,不同生活型植物的日平均光合速率、叶片氮、磷的组成均有所不同。3种乔木的日平均光合速率、叶片氮含量、叶片磷含量及叶片氮磷含量比分别在2.36~10.72 μmol/(m2·s),17.2~19.87 mg/g,1.66~6.9 mg/g,2.49~11.25之间,4个指标中,竹柳相对较高,柑橘相对较低。4种灌木日平均光合速率、叶片氮含量、叶片磷含量及叶片氮磷含量比分别在4.71~10.46 μmol/(m2·s),22.06~28.61 mg/g,2.05~5.27 mg/g,4.18~13.95之间,4个指标中,地挑花相对较高,苎麻相对较低。8种草本日平均光合速率、叶片氮含量、叶片磷含量及叶片氮磷含量比分别在4.16~11.11 μmol/(m2·s),9.18~38.76 mg/g,1.47~9.5 mg/g,2.96~10.15之间,4个指标中,鬼针草相对较高,风车草相对较低。
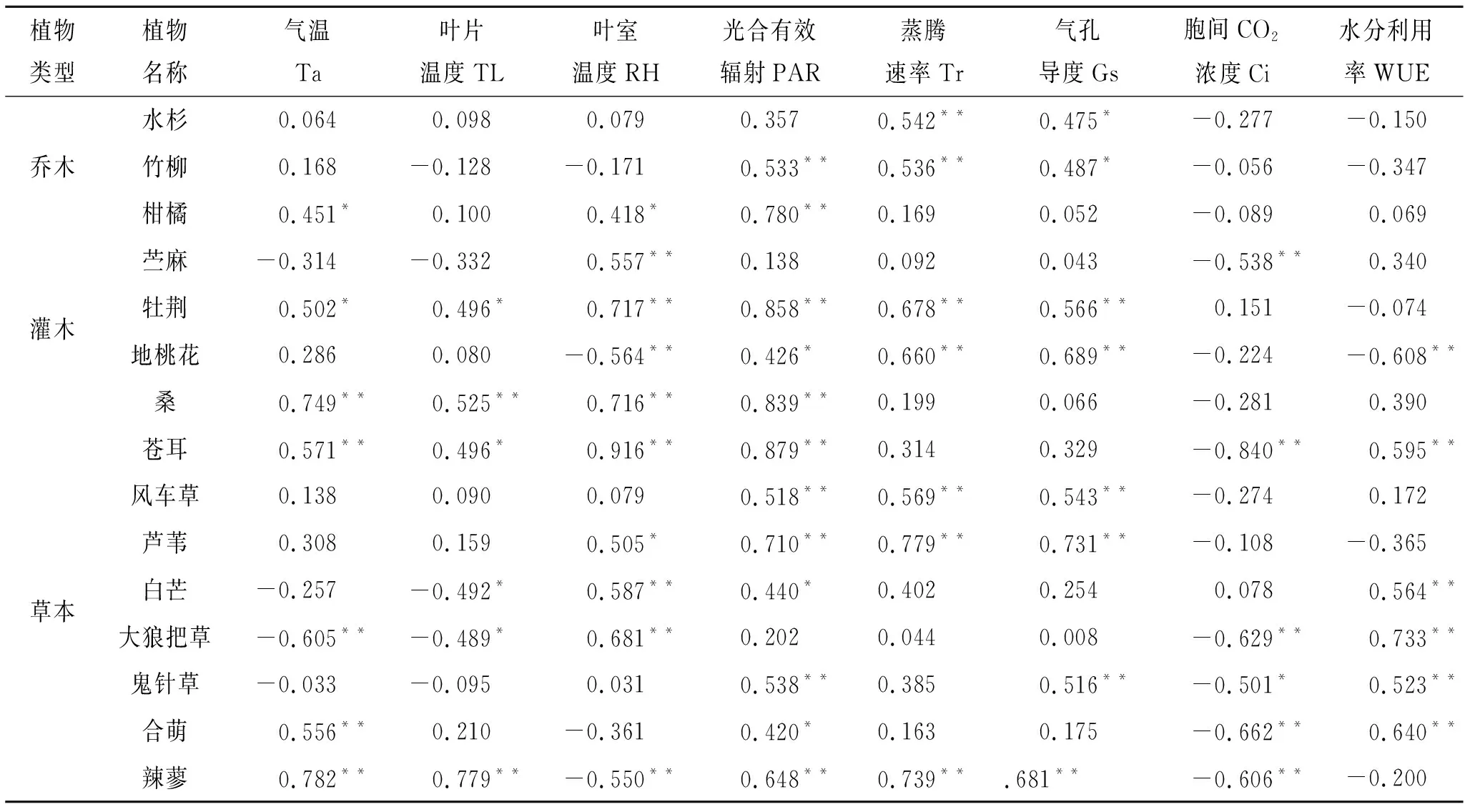
表1 不同树种净光合速率与影响因子相关性

表2 植物的日平均光合速率和叶片氮、磷含量计量学特征
3.2.2 植物日平均光合速率与叶片氮、磷含量的相关性分析 植物叶片细胞中氮磷浓度及氮磷比与光合速率密切相关[17]。由表3分析可知,不同生活型植物叶片日平均光合速率与叶片全氮、全磷及氮磷比之间的关系存在差异。乔木叶片和草本叶片的日平均光合速率与叶片全氮、全磷及氮磷比之间相关性均不显著,而灌木叶片的日平均光合速率与叶片全氮、全磷的相关性均不显著,与氮磷比之间呈显著负相关。
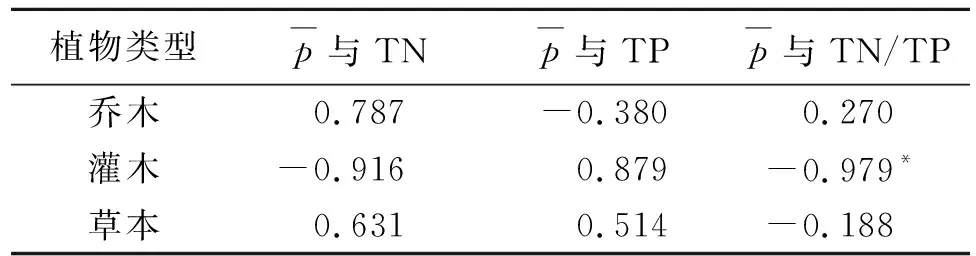
表3 不同生活型日平均光合速率与叶片氮、
3.3 固碳释氧能力分析
净同化量是植物单位时间内光合作用产生的有机物与呼吸作用消耗的有机物之差。净同化量越大,说明植物的生产力越强,有机物累积量越大,固碳能力越大[18]。由图1可知,不同类型植物日光合净同化量、单位叶面积固碳量及单位土地面积固碳量均不相同,但同一植物日光合净同化量、单位叶面积固碳量变化趋势相同,而单位土地面积植物固碳量受叶面积指数的影响,变化趋势略有差异,乔木中,竹柳的日光合净同化、单位叶面积固碳量及单位土地面积固碳量均最高,分别为:405.17 mmol/(m2·d),14.26 g/(m2·d),64.46 g/(m2·d);灌木中,牡荆的日光合净同化、单位叶面积固碳量及单位土地面积固碳量相对较高,分别为:282.68 mmol/(m2·d),9.95 g/(m2·d),65.47 g/(m2·d),其次为地桃花。草本中,鬼针草、合萌相较具有较高的日光合净同化、单位叶面积固碳量,分别为:410.20 mmol/(m2·d)和14.44 g/(m2·d),406.63 mmol/(m2·d)和14.31 g/(m2·d),受叶面积指数影响,芦苇和辣蓼日光合净同化、单位叶面积固碳量虽较鬼针草、合萌低,但其叶面积指数相对较高,其单位土地面积固碳量相对较高,分别为:57.51 g/(m2·d),55.54 g/(m2·d)。
4 讨论与结论
(1) 植物的光合作用是非常复杂的内部生理过程,净光合速率的大小通常作为反映了植物固碳释氧能力的强弱[19],评价内因与外因对光合作用影响程度的指标[20],而净光合速率受外界环境因子影响较大。在净光合速率与外界多变的环境因素与植物生理因素相关性分析中,测试的15种植物中,苎麻、白芒、大狼把草及鬼针草的净光合速率则与气温、叶片温度,叶室湿度以及水分利用呈负相关,其中白芒及大狼把草净光合速率则与气温、叶片温度呈显著(p<0.05)或(p<0.01)极显著的负相关,这与邵永昌[21]、王海湘[22]等的研究结果不同,究其原因,可能因为测定时间段内温度过高减弱了植物光合碳同化过程。试验所测的大多数植物的净光合速率与光合有效辐射、蒸腾速率及气孔导度等呈显著(p<0.05)或(p<0.01)极显著的正相关,仅少数种类植物的净光合速率与气孔导度无关,可能由于测定时间段内温度过高,植物处于高温胁迫状态,使得其非气孔限制因子对光合作用的影响占主导地位[23];而测试植物的日平均光合速率与胞间CO2浓度呈显著(p<0.05)或(p<0.01)极显著的负相关,这与邵永昌[21]、吴芹[24]的研究结果一致。说明一定程度内,光合有效辐射、气孔导度、蒸腾速率的变化及胞间CO2浓度是影响植物叶片光合速率大小的主要因素,从而影响其固碳能力。故而在开展植物生态修复工程中,应保持适当种植距离,以增强植被采光及通风效果,从而提高植物的固碳能力。

注:P为测定日单位叶面积的净同化总量;WCO2为测定日单位叶面积的净固定CO2的质量;QCO2 为测定日单位土地面积的净固定CO2的质量.
单位土地面积固碳量
(2) 消落带湿地系统作为物质转换的重要动态中心,其他元素的输入和转化对碳循环动态也产生着重要影响[4],尤其以氮磷对植物光合固碳能力及碳分配有着显著的影响。试验测得的不同生活型的植被的全氮含量平均值分别为:18.6 mg/g,24.59 mg/g,22.05 mg/g,高于三峡库首地区的氮含量(18.3 mg/g),与同尺度区域的植物叶片氮素含量相当[25-27];不同生活型的植被的全磷含量平均值分别为3.55 mg/g,3.15 mg/g,4.02 mg/g,而磷含量要高于其他同尺度区域[26,28],这表明,重庆三峡库区消落带植物体内氮磷含量较为富足。同时,试验测得的不同生活型的植被的日平均光合速率均高于其他地区[29]。此结果可以通过以下假说来解释:在淹水生境下维持相对高的光合速率以及叶片营养元素对于植物存活是非常关键的[30]。通过对不同生活型植物的日平均光合速率与叶片全氮、全磷之间相关性分析结果显示两者显著性并不明显,而在与叶片氮磷比相关性方面,灌木的日平均光合速率与叶片氮磷比呈显著负相关,可能因为在库区消落带氮磷供给充足的前提下,叶片磷供给量过多时,使得地上部分生物量积累受到限制[31];同时,也可能与土壤环境有关,土壤对植物的叶片氮磷含量变化有显著影响,本研究未就土壤因子对植物叶片氮磷含量的影响进行研究,或许会忽略土壤环境对植物固碳功能的限制影响[32],需继续深入开展研究。
(3) 固碳能力是反映植物光合作用的重要指标。净固碳量高说明植物与外界交换的CO2和O2较多,可以把更多的太阳能转化成有机质储存于植物体内[33],日光合净同化量、单位叶面积固碳量及单位土地面积固碳量均不相同。测试的15种植物中,乔木中的竹柳,灌木中牡荆、地桃花,具有较高的日光合净同化量、单位叶面积固碳量及单位土地面积固碳量,由于净光合速率和叶面积指数是决定单位土地面积植物固碳量的重要因素[34],草本中的鬼针草、合萌虽然具有较高的日光合净同化量、单位叶面积固碳量,因为其叶面积指数远远低于其他植物,草本中芦苇和辣蓼等单位土地面积的固碳能力较其他植物要高。因此,本研究建议在三峡库区消落带植物修复进行植被选择过程中,在充分考虑植物的固碳的生态效益时,应选择诸如竹柳,牡荆,地桃花,芦苇,辣蓼,鬼针草,合萌等高效碳汇植物,在三峡库区消落带生态修复工程进行植被选择配植过程中,合理搭配,构建多层复合配置的林灌草的模式[35],不仅提高整个植被系统的固碳能力,从而提高库区消落带湿地生态系统的碳汇功能。
三峡库区消落带湿地生态系统受水文机制调控,海拔变化引起的淹水持续时间、频率和水深对其原有生态系统平衡造成严重破坏,植物群落的组成与空间分布格局遭受显著影响[36-39]。库区消落带的植被修复措施用来改善消落带生态系统逐渐成为人们关注的焦点[40]。光合固碳作用是植物的重要生态功能之一,对于维持消落带湿地生态系统平衡具有重要的价值。而植物的光合固碳过程是一个复杂的生理过程,受到外界多变的环境因素的影响。在今后的研究中,应结合多种分析方法将这些相互联系、相互制约的影响因素综合起来,筛选具有较强固碳能力及碳储量潜能的植物类型,并考虑植物的生长适应性来优化植物配置模式。
猜你喜欢
今日农业(2022年14期)2022-11-10
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
现代农业科技(2016年22期)2017-03-24
学周刊·下旬刊(2015年10期)2015-07-22
中国烟草学报(2012年1期)2012-04-09
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2010年3期)2010-11-16
