
狭果秤锤树自然种群分布格局
2020-04-22 05:42周赛霞高浦新潘福兴胡余楠刘向平彭焱松
浙江农林大学学报 2020年2期
周赛霞,高浦新,潘福兴,龙 晶,胡余楠,刘向平,彭焱松
(1.江西省-中国科学院庐山植物园 江西省植物迁地保护与利用重点实验室,江西 庐山 332900;2.江西省寻乌县林业局项山林业工作站,江西 寻乌342200)
近年来,分布格局研究成为生态学研究领域的热点问题[1-3]。自然状态下,各类植物种群通常并非均匀分布,而是依不同年龄、不同研究尺度呈斑块状分布,形成复杂的种群空间分布特征。这些分布特征是对种群自身生物学特性和外在生态环境的一种表达,并且反过来影响着种群的发展趋势[4]。种群的空间分布格局可以反映物种在群落中不同尺度下的分布类型或配置方式,同一物种在不同的生活史阶段、不同的生境条件也可能表现出完全不同的分布类型[5-7]。而且分布格局具有尺度依赖性,在一个尺度表现为聚集分布,在其他尺度则可能表现为均匀分布或随机分布[8]。一般来说,如果竞争对种群起着主要制约因素,种群趋向于均匀分布;如果生物学特性起主要作用,则大多为聚集分布;如果以上两者都不起决定性作用,种群一般表现为随机分布[4]。种内关联性可以分析种群不同生长阶段个体间的功能关系和相互作用[9]。空间正关联表明相互促进及互利的生态关系,空间负关联表明相互排斥及不利的生态关系,空间无关联则意味着个体间无明确的生态关系[10-11]。狭果秤锤树Sinojackia rehderiana为安息香科Styracaceae秤锤树属Sinojackia植物,仅间断分布于华南和华东,分布范围狭窄且个体数量少。狭果秤锤树性喜湿润的沟谷常绿阔叶林内,分布海拔较低,易受到人为干扰。生境破坏和自身种子萌发率低等原因导致分布范围逐渐变窄[12],有标本采集记录的有些产地已找不到该种的分布。目前,已有一些学者对其群落区系[13]、 年龄结构和分布格局[14]、 群落结构及更新[15]、 种间联结性[16]、 遗传印记[17]等方面做过研究,而关于种群空间格局和关联性研究则鲜有报道。本研究旨在为该群落和狭果秤锤树种群就地保护提供理论支持,为迁地保护种群空间配置提供建议。
1 研究区域和方法
1.1 研究区域概况
研究区地处江西省九江市永修县青山村村前一处风水林(29°07′55.8″~29°08′15.5″N, 115°42′39.8″~115°42′52.7″E),海拔高度约23~25 m。林地面积约7 hm2,地势平坦,内有一条狭长的河沟,外围由田地和乡村小道环绕,部分地段人为活动频繁。该地属亚热带湿润季风气候区,年平均气温17.5℃,最冷月1月平均气温5.0℃,最热月7月平均气温29.1℃,年均降水量1 560.9 mm,年均相对湿度78%。土壤为沙质壤土,pH 6.0~6.5。该风水林为亚热带常绿阔叶林,垂直结构明显,乔木上层以樟树Cinnamomum camphora、紫弹树Celtis biondii占绝对优势,乔木下层以狭果秤锤树、尾叶冬青Ilex wilsonii和瓜木Alangium platanifolium为优势种;灌木层分布最多的为狭果秤锤树和瓜木、大青Clerodendrum cyrtophyllum等;草本层主要物种为江南短肠蕨Allantodia metteniana、接骨草Sambucus javanica、三脉紫菀Aster trinervius subsp.ageratoides等。
1.2 研究方法
1.2.1 样地设置和调查 采用相邻格子样方法,在人为活动相对较少,狭果秤锤树集中分布地段,设置1个面积50 m×100 m的固定监测大样方,调查时将大样方分成50个10 m×10 m的小样方,小样方四角用罗盘仪定位,安装PVC-U管,距离用皮尺测量。采用每木调查法对小样方内所有狭果秤锤树进行统计,记录每株植物的胸径(地径)、树高、冠幅以及相对位置坐标、生长状况等信息。在1.3 m高度以上有萌枝的个体只记录主干信息,若萌枝在1.3 m以下分别记录各枝信息。以大样方西南角为坐标原点,东西向为坐标横轴,南北向为坐标纵轴,记录植株坐标位置。
1.2.2 种群径级和高度级结构 在相同的环境下,种群的年龄结构与径级结构对环境具有一致的反应规律[18]。本研究采用以径级代替年龄级的方法对狭果秤锤树进行径级的划分,胸径(DBH)以1 cm为一级,即0<DBH<1 cm为第1级,1≤DBH<2 cm为第2级,依此类推。对狭果秤锤树高度级(H)的划分,以1 m为一级,方法同径级划分。将狭果秤锤树按径级和高度级,分别统计株数,采用SigmaPlot 14.0软件制表,绘制种群结构柱状图。
根据狭果秤锤树生活史以及种群实际年龄大小情况,将狭果秤锤树按不同的大小级分成4个生活史阶段,定义0<DBH<1.0 cm的植株为幼苗、1.0≤DBH<2.5 cm的植株为幼树、2.5≤DBH<6.0 cm植株为中树、DBH≥6.0 cm植株为成树。
1.2.3 点格局分析 分别对狭果秤锤树不同生活史阶段进行分布格局和类间关联性点格局分析。对某一生活史阶段进行空间分布格局分析时,采用单变量完全空间随机模型(CSR),将空间尺度限定在半径小于10 m内。根据Monte Carlo模拟结果,单变量O(r)值在置信区间之上为该尺度上的聚集分布,O(r)值大小代表聚集强度大小;O(r)值在置信区间之内则为该尺度下的随机分布;O(r)值在置信区间之下为该尺度下的均匀分布。不同生活史阶段进行点格局双变量分析,如果双变量O(r)值分别在置信区间上、置信区间内、置信区间下时,分别表明两者在该尺度空间正相关、相互独立、负相关[19-22]。
点格局分析采用生态学软件Programita(2014),研究尺度0~25 m,设定圆环宽度为1 m。运用Monte Carlo循环199次,95%置信区间。
2 结果与分析
2.1 狭果秤锤树径级结构与高度结构
狭果秤锤树径级结构见图1。样地内共调查到狭果秤锤树1 275株,胸径最大为15.7 cm。径级1~16的株数分别为321、 534、 159、106、 45、 46、27、 13、 9、 6、 2、 2、 2、 1、 0、 2株。 种群径级结构完整,小径级个体多,幼苗、幼树充足,表明种群更新能力较强;大径级个体相对较少,体现了狭果秤锤树灌木或小乔木的生物学特征。种群高度级结构见图1。高度级可表现种群在垂直空间上的配置状况[23],与径级结构结合可更好地表现种群的大小状态。狭果秤锤树高度级完整无缺失,与径级结构类似,低矮个体较多,其中1~2 m高个体数量最多;5 m以上个体数234株,最高个体11 m。径级结构和高度级结构表明狭果秤锤树种群发展稳定,正处于增长阶段。
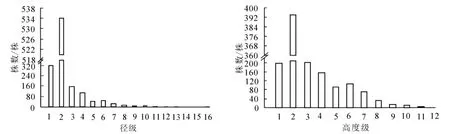
图1 狭果秤锤树种群径级结构和高度级结构Figure 1 DBH classes and height classes of S.rehderiana population
2.2 狭果秤锤树种群不同生活史阶段个体分布状况
狭果秤锤树幼苗321株、幼树639株、中树251株、成树64株。各生活史阶段个体均散生在样地中(图2),其中样地正西北中线附近个体较少,样地东北角分布有较多的幼苗和幼树,中树和成树分布的区域均有幼苗和幼树分布。
2.3 狭果秤锤树种群不同生活史阶段分布格局
根据图3可初步判断,狭果秤锤树幼苗在0~21 m的尺度上呈显著聚集分布,21~25 m的较大尺度上呈随机分布;幼树在0~11 m的尺度上聚集分布,18~20 m的尺度上均匀分布,其他尺度上随机分布;中树在0~4 m和7~8 m的较小尺度上聚集分布,其他尺度上随机分布;成树在0~1 m和2~4 m的较小尺度上聚集分布,其他更多尺度上表现为随机分布。总之,随着龄级的增加,狭果秤锤树在更多的尺度上表现出随机分布,分布格局表现为聚集分布向随机分布过渡的趋势。
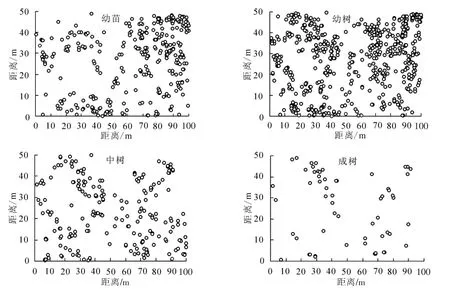
图2 狭果秤锤树不同发育阶段个体在样地中的分布点图Figure 2 Distribution of individuals of different development stages of S.rehderiana population

图3 狭果秤锤树不同生活史阶段个体点格局分析Figure 3 Spatial distribution pattern of individuals at different life stages of S.rehderiana population
2.4 狭果秤锤树种群不同生活史阶段空间关联性
幼苗与幼树小尺度上空间正相关,在其他更大更多的尺度上无明显空间相关性;幼苗与中树在全部尺度上表现出空间负相关与无明显相关性间断出现的趋势;幼苗和成树之间在大部分尺度上无明显空间相关性,仅在14.5~17.5 m尺度上空间负相关;幼树与中树在0~4.5 m尺度上表现为正相关,13.5~25.0 m尺度上负相关,其他尺度无明显相关性;幼树与成树在较小的尺度0~14.5 m空间正相关或不相关,14.5~25.0 m尺度上空间负相关;中树与成树在大部分尺度上空间正相关或不相关,仅在17.5~19.5、20.5~21.5 m尺度上表现出负的相关性(图4)。总之,种群负相关性主要表现在幼苗与中树、幼树与中树、幼树与成树间,且都为较大尺度上。
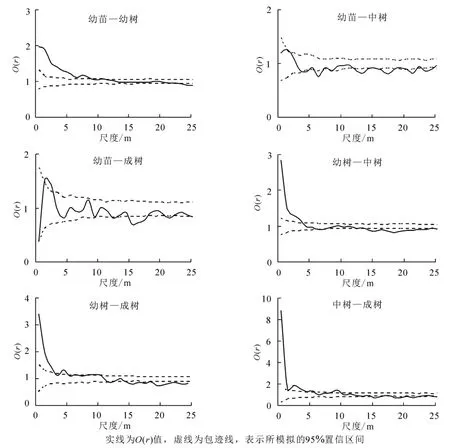
图4 狭果秤锤树不同生活史阶段个体空间关联性Figure 4 Spatial relationship of individuals at different life stages of S.rehderiana population
3 结论与讨论
种群的径级结构和高度级结构不仅能反应种群的更新类型,而且能推测种群未来的发展趋势[23]。狭果秤锤树种群径级结构和高度级结构表现出一致的规律,小径级或较矮个体多,大径级或较高个体少,种群表现出稳定增长的趋势。这可能与种群所处的自然环境密不可分。调查区域地带性植被为常绿阔叶林,上层高大乔木枝繁叶茂,林内一条很长的河沟,造成林内小环境比较阴湿,而且土壤为砂性壤土,透气性很好且不积水,不会造成土壤板结和水分过饱和,天然适合于果皮坚硬且具有深休眠特性[16]的狭果秤锤树种子沙藏,导致样地内狭果秤锤树实生苗丰富。另外,样地内萌孽产生大量的萌生苗,种群表现为小个体较多,更新状态良好。
种群不同生活史阶段表现出不同的分布格局类型,这样不同年龄级的个体可以交错利用有限的环境资源[22-23]。许多研究表明:物种自身的生物学特性、其他物种的影响或者环境因素对种群空间格局均会造成影响[24-25]。狭果秤锤树不同生活史阶段个体均在小尺度上表现为聚集分布,且随着尺度的增大,聚集程度逐渐减弱;随着龄级的增加,逐渐表现出更多尺度上的均匀分布或随机分布。其原因可能是:狭果秤锤树结实量大,且种子较重,扩散能力差,掉落后主要分布在母株周围;研究区域环境适合狭果秤锤树种子萌发,产生大量的实生苗,且物种本身萌孽能力较强,产生大量的萌生植株,致使种群在较小的尺度上聚集分布;二是聚集分布更有利于形成适合种群自身生长的小环境,可以更好地发挥群体效应,以增强种间竞争力,抵御其他物种入侵,更好地维持种群的长远发展[7];三是随着龄级的增加,个体对环境中有限的资源竞争加剧,导致种内出现激烈的竞争,自疏和它疏作用增强,部分林木死亡,成熟个体向均匀分布或随机分布方向发展。
许多研究表明种群各龄级之间的相关性受龄级个体大小的影响,龄级差异越大,个体间正相关性越弱,更多的表现为不相关或负相关性[10,19,26-27]。形体较大的个体可能对形体较小的个体产生不利的影响,可以理解为与种内资源竞争有关,与本研究的结论相一致。狭果秤锤树空间负相关性主要表现在幼苗与中树间、幼树与中树间、幼树与成树间,反映了大径级个体对其幼苗幼树的生长有强烈的抑制作用;狭果秤锤树幼苗与幼树因形体较小,对土壤养分、光照等环境资源的竞争力较弱,在长期的生长过程中相互庇护,表现出明显的正相关关系;随着幼龄个体的不断生长,个体对资源的需求量加大,个体间相互有利的关系被打破,表现出相互胁迫的竞争关系,从而出现中树或成树对幼苗和幼树的不相关或负相关关系。
自然状态下,狭果秤锤树的结构和空间分布格局受其生物学特性和环境因素的综合影响,种群的空间格局和关联性研究仅是分析群落空间格局与过程的第一步[20],群落内狭果秤锤树种群与其他种群的空间关系,样地内各种环境因子与种群空间分布格局的相互作用如何尚不明确,均有待于深入研究。狭果秤锤树在华东和华南分布区域的缩减反应了适合的生态环境和植被的减少,在人类干扰频繁地区,如何减少对森林植被的影响和破坏,可持续的营林措施和管理决策显得至关重要。自然分布于永修县的这片狭果秤锤树林天然更新良好,在就地保护过程中要尽量避免人为干扰破坏,杜绝进林中取土挖坑、砍伐等活动;在迁地保护过程中努力营造类似原生境的立地条件,特别是土壤条件,提高苗木成活率和生长速度,促进狭果秤锤树种群的天然更新。
猜你喜欢
北华大学学报(自然科学版)(2022年5期)2022-11-21
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
伦理学研究(2021年6期)2021-12-29
园艺与种苗(2021年3期)2021-05-26
新农民(2020年2期)2020-12-08
伦理学研究(2020年4期)2020-09-07
河北果树(2020年1期)2020-02-09
种子(2019年6期)2019-07-13
农民致富之友(2016年24期)2017-04-26
