
陆地棉小GTP结合蛋白基因GhRop4的克隆及其表达分析
2020-04-22 08:21李月吾尕力汗阿不都维力周垚均刘超任艳萍郭旺珍刘晓东
棉花学报 2020年1期
李月,吾尕力汗·阿不都维力,周垚均,刘超,任艳萍,郭旺珍,刘晓东*
(1.新疆农业大学/ 农业生物技术重点实验室,乌鲁木齐830052;2.南京农业大学/ 作物遗传与种质创新国家重点实验室,南京210095)
作为1 类分子量为20~30 kD 的结合蛋白,小 GTP(Guanosine triphosphate,三磷酸鸟苷)结合蛋白是许多信号转导过程的中心调节器,广泛存在于真核生物中[1-2]。 根据小GTP 结合蛋白的结构和功能分为 Ras、Ran、Rab、Rho 和 Arf 5 个超家族[1-3]。 在真核生物中,Ras 和 Rho 家族蛋白是信号转导开关,而其它3 个家族蛋白的功能主要参与囊泡和大分子运动的调节[1-2]。 然而,高等植物有1 个独特的Rho 亚家族的小GTP 结合蛋白称为 Rop 蛋白[4-5]。 Rop 蛋白具有 GTP 酶活性,与 GTP 结合可以催化 GTP 水解, 而与 GDP(Guanosine diphosphate,二磷酸鸟苷)结合无催化活性,通过与GTP/GDP 循环结合,在植物信号传导中作为分子开关调控相关代谢活动[6]。 它由N末端催化结构域G 结构域 (Catalytic G-domain)和 C 末端高变区 (Hyper-variable domain,HVR)组成, 其中G 结构域负责与核苷酸及效应子结合,HVR 负责亚细胞靶向。 G 结构域含有5 个保守序列基序称为 G-box 基序(Ⅰ-Ⅴ),这5 个基序是核苷酸结合、GTP 水解和Mg2+结合的功能区域。 Ⅰ、Ⅲ是GTP 酶结构域,Ⅱ和Ⅲ基序也被称为开关Ⅰ(SwitchⅠ)和开关Ⅱ(SwitchⅡ),Ⅲ~Ⅴ是GTP 或GDP 的结合区域这些区域在与GDP 和GTP 结合状态下具有不同的构象, 对与效应蛋白和调节蛋白的短暂相互作用至关重要。G 结构域内还包含1 个Rho 族独有的螺旋域,称为插入域[5]。 Winge 等[7]根据 Rop 蛋白 C 端氨基酸序列差异, 将Rop 家族蛋白分为2 种类型,即Ⅰ型和Ⅱ型。 Ⅰ型 Rop 蛋白 C 端以典型的CaaL 盒基序终止,其中 C 为半胱氨酸(Cys),a 为脂肪族氨基酸残基,L 为亮氨酸 (Leu),Cys 残基被C20 异戊二烯基脂香叶基香叶基修饰。 Ⅱ型Rop 以 GC-CG 盒终止 (其中 G 为甘氨酸 Gly,C是 Cys),其中 2 个 Cys 残基是由 5 或 6 个脂肪族残基隔开并经C16 棕榈酸酯或C18 硬脂酸酯脂肪酸 S 酰化[5]。
自从第1 个Rop基因在豌豆中被鉴定出来,目前已经在多种植物中鉴定出Rop基因,其中拟南芥中有 11 个、玉米 9 个、油菜 11 个、葡萄 7个、水稻7 个、苜蓿7 个、烟草6 个、巴西橡胶树5个、番茄 9 个[8-9]。 根据对Rop基因的功能研究,Rop 家族蛋白具有多种功能,包括调控花粉管尖生长、植物细胞生长、信号传导、核迁移和细胞骨架形成等[10-12]。 除此之外,还广泛参与环境应激反应和宿主- 病原体相互作用等过程[13]。 在寒冷条件下,苹果的Rop基因转录表达增加,导致果实中乙烯和活性氧含量降低[14]。 拟南芥AtRop11 的表达影响种子萌发、幼苗生长、气孔关闭、脱落酸介导响应和干旱胁迫响应[15]。 在烟草中过量表达NtRop1 基因,转基因植株产生较多的H2O2,对盐胁迫的耐受能力降低[16]。将苜蓿Rop基因MfARL1转入拟南芥, 转基因植株的Na+/K+比低于对照株,并增强了盐胁迫的耐受性[17]。 拟南芥Rop 效应蛋白(RIC1)敲除植株在盐胁迫下的存活率增加[18]。 在水稻中发现7 个功能各异的Rop基因,OsRac1 通过调控活性氧爆发和细胞死亡, 正调控水稻对稻瘟病的抗性;OsRac4 和OsRac5 负调控水稻稻瘟病抗性,其它基因可能与水稻稻瘟病无关[19]。 烟草NtRac5 通过调控烟草细胞氧化酶参与烟草抗病反应中的活性氧爆发[20]。MtRop9参与植物受病原菌侵染的早期抗性反应[21]。HvRacB参与大麦对白粉病菌的基础防卫反应[22]。棉花中Rop家族基因的研究报道主要和棉花纤维发育和陆地棉黄萎病菌侵染反应有关。 李先碧等[23]从陆地棉克隆了2 个棉花纤维起始和伸长时期优势表达的GhRacA和GhRacB,推测可能调控棉花纤维早期发育。 Deborah 等[24]克隆的GhRac9、GhRac13 基因,在棉纤维发育过程中初生和次生壁合成的过渡时期的优势表达,推测它们可能是通过产生H2O2介导棉花次生壁的合成。 GhRac1调控细胞骨架的构建是纤维伸长的潜在调节因子[25]。 王钰静[26]利用病毒诱导基因沉默(Virusinduced gene silencing, VIGS) 技 术 抑 制 棉 花GhRop6 基因的表达, 沉默植株的木质素含量较对照组低,并产生较多的H2O2,推测GhRop6 基因正调控棉花对黄萎病的抗性。 而关于棉花中Rop基因在高盐、干旱、低温等逆境条件下的表达谱鲜有报道。
棉花是重要的纤维和油料作物,具有重要的经济作用,因此被广泛种植。 但是棉花在生长过程中往往受到各种生物胁迫和环境因素如害虫、病原体、盐、冷和干旱等的影响。 这些不利因素对棉花的生长、产量以及纤维品质影响很大,造成巨大的经济损失。植物特有的Rop 家族蛋白作为上游信号传递的分子开关,参与植物抗逆反应和对病原菌的基础防卫调控。 本文利用生物信息学的方法,从陆地棉中克隆1 个Rop基因GhRop4,对其蛋白序列特征、逆境和组织表达特性进行研究,为进一步揭示棉花GhRop4 基因的生物学功能提供重要信息。
1 材料与方法
1.1 材料
陆地棉抗旱材料KK1543 和黄萎病菌菌株V991 均由新疆农业大学农学院农业生物技术重点实验室提供。
1.2 方法
1.2.1播种、生长条件和胁迫处理。将KK1543种子均匀播种于黑土和蛭石2∶1 混合的营养土中,在温度(28±2)℃、湿度 65%、光照 / 黑暗周期为16 h/8 h 条件下连续培养15 d。 选取生长一致的棉花,置于吸水纸上吸干水,分别进行高盐(200 mol·L-1NaCl)、干旱、4 ℃低温处理,具体处理参照李月等[27]已报道的方法进行。 分别于处理后 0 h、1 h、3 h、6 h 和 12 h 采集叶片,同时采集未处理材料的根、真叶、子叶、下胚轴和茎用于组织特异性表达检测。 将以上各时间点采集的不同组织的材料迅速置于液氮速冻,用于总RNA 提取,进行目的基因表达分析。
将 KK1543 种子在 30% H2O2溶液中浸泡2 h,无菌水冲洗3 次,最后在无菌水中避光浸泡12 h, 将露白的棉种平铺在发芽纸上发芽2 d,选取萌发一致的种子播种于黑土和蛭石2∶1 混合的营养土中,生长条件同1.2.1。 待2 片真叶展平时,采取伤根蘸菌法[26],进行黄萎病菌V991 侵染。 在接菌 0 h、4 h、8 h、12 h、24 h 和 48 h 后取棉苗的幼根,迅速置于液氮冷冻,用于总RNA 提取及基因的表达分析。
1.2.2总RNA 的提取和cDNA 的制备。采用杭州博日科技公司的BSC65S1 Biospin 多糖多酚植物总RNA 提取试剂盒提取棉花RNA, 赛默飞(Thermo fisher Scientific)DNase I 去除 RNA 中残存 的 DNA, 利 用 Thermo Scientific(NanoDrop 1000) 核酸分析仪和1.2%琼脂糖凝胶电泳检测RNA 质量和完整性,质量检测合格的RNA 按照北京全式金生物技术有限公司 TransScript○RII First-Strand cDNA Synthesis SuperMix 反转录试剂盒说明书合成cDNA,用于基因表达分析。
1.2.3棉花GhRop4 基因的克隆。以拟南芥TAIR (http://www.arabidopsis.org/index.jsp)的AtRac1 基因(AT2G17800)的序列作为 BlastP 查询序列, 在棉花EST 数据库(http://www.leonxie.com),棉花基因组数据库(http://www.phytozome.net/)中比对,获得了 1 个 cDNA 序列,然后根据此序列的ORF(Open reading frame)序列设计引物GhRop4F:5’-ATGAGCGCCTCGAGATTCATA-3’和 GhRop4R:5’-TCATAATATTGAGCAACCACC-3’。 以陆地棉 KK1543 的 cDNA 为模板,以高保真聚合酶TransStar KD Plus(北京全式金)进行PCR 扩增。将扩增产物经1%琼脂糖电泳检测,回收目的片段连接至peasy-Blunt Zero 克隆载体,进行质粒酶切鉴定,将阳性克隆质粒送昆泰锐生物技术有限公司测序验证。 参照李月等[27]方法进行试验所用的PCR 程序、扩增产物检测及目的片段的纯化回收。
1.2.4序列分析和聚类分析。利用NCBI 在线服务工具中BlastP 和DNAMAN 软件对GhRop4 蛋白序列进行氨基酸同源序列分析和多重序列比对。同时在NCBI 上查找水稻Oryza sativaOsRac1(XP_015621645.1)、OsRac3 (XP_015625155.1)、OsRac4 ( XP_015641323 .1 ) 和 OsRac5 ( XP_015627011.1); 拟 南 芥Arabidopsis thalianaAt-Rop1 (NP_190698.1)、AtRop2(NP_173437.1)、AtRop4 (NP_177712.1)、AtRop5(NP_195320.1)、AtRop6(NP_ 001190917.1)和 AtRop11(NP_201093.1); 小 麦Triticum aestivumTaRop3(ADD23345); 烟 草Nicotiana tabacumNtRac1(XP_016513031.1)、NtRac5 (XP_016512162.1);玉米Zea maysZmRop1(XP_008644256.1);大麦Hordeum vulgareHvRac1(CAD57743.1)、HvRac3(CAD57742.1)、HvRacB(CAC83043.2)和 HvRop4(CAD27896.1)6 个物种功能已知的 Rop 蛋白序列。 利用Clustal X 和MEGA 5 软件采用邻接法方法对棉花GhRop4 和上述物种的Rop 蛋白序列构建系统发生树, 分支的可靠性评价采用靴带分析。
1.2.5棉花GhRop4 基因的表达分析。利用多糖多酚植物总RNA 试剂盒提取棉花不同处理、不同时间点棉花RNA 和棉花不同组织的RNA,质量检测合格反转录获得cDNA 第一条链,以此链为模板进行荧光定量PCR 扩增, 扩增体系和反应条件参照李月等[28]的方法。 内参Ubiquitin7 基因引物序列为 UBQ7F:5′-CCAGAAGGAATCCACTTTGC-3′;UBQ7R:5′-CCAGCTCACATCAGCATACG-3′ 和GhRop4 基 因 的 引 物 序 列qGhRop4F:5′-GGTCTTCTGCCTACATCGAGT-3′;qGhRop4R:5′-GAAATGGGCACTGCGTTAGG-3′。 采用 2-△△CT法分析数据,确定基因的相对表达量。并利用DPS 7.05 软件对定量数据进行单因素方差分析。
2 结果与分析
2.1 GhRop4基因的克隆与序列分析
以 KK1543 叶片 cDNA 为模板进行 PCR 扩增结果(图1)表明,从陆地棉中克隆1 个Rop基因 cDNA 序列, 命名为GhRop4, 基因登陆号AY965614.1。GhRop4 基因的 ORF 为 588 bp,编码195 个氨基酸,分子量为21.229 kD,理论等电点为8.81,为碱性蛋白。 利用DNAMAN 软件与拟南芥、水稻、大麦3 个物种中5 个ROP 蛋白序列比对发现,GhRop4 蛋白符合植物Rop 蛋白的特征,含有5 个(Ⅰ-Ⅴ)效应结构区Ⅰ、Ⅲ是GTP酶结构域,Ⅱ是效应因子结构域,Ⅳ、Ⅴ是GTP或GDP 的结合区域,2 个开关区域 (开关Ⅰ和开关Ⅱ),1 个Rop 插入序列及C 末端高度变异区(HVR)(图 2)。Rop 插入序列包含 10 个氨基酸残基(DQQFLTDHFN)。 GhRop4 蛋白 HVR 区的氨基酸序列C 末端为CSIL。

图1 GhRop4 基因的克隆Fig.1 Cloning of GhRop4 gene
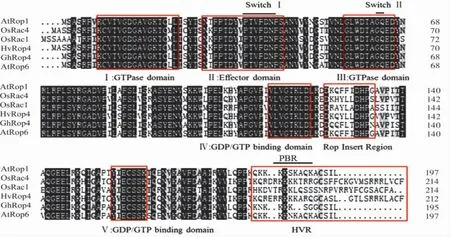
图2 GhRop4 功能结构域分析Fig.2 Analysis of the functional domains in GhRop4 protein
2.2 棉花GhRop4蛋白系统进化树分析
Winge 等[7]根据 Rop 蛋白 C 端氨基酸序列差异,将Rop 家族蛋白分为2 种类型,即Ⅰ型和Ⅱ型。从GenBank 中下载水稻、拟南芥、小麦、烟草、玉米、大麦中共19 个功能已知Rop 蛋白序列,构建系统进化树。 分析表明(图 3),19 个 Rop 蛋白分为 2 类,12 个 Rop 蛋白聚为Ⅰ型 Rop 蛋白,7个Rop 蛋白聚为Ⅱ型 Rop 蛋白,GhRop4 蛋白属于Ⅰ型Rop 蛋白。 研究表明,Ⅰ型Rop 蛋白主要参与调控植物生长发育、植物抗逆和抗病性反应,推测属于Ⅰ型Rop 蛋白的棉花GhRop4 蛋白在植物功能反应中扮演多样角色。 利用DNAMAN 软件对这 6 个已克隆的GhRop基因(GhRacA、GhRacB、GhRac9、GhRac13、GhRac1、GhRop6)与本研究克隆的GhRop4 基因进一步比对分析,GhRop4 基因与GhRac1 基因的序列相似性达到88%, 与其它5 个GhRop基因的相似性均大于75%。
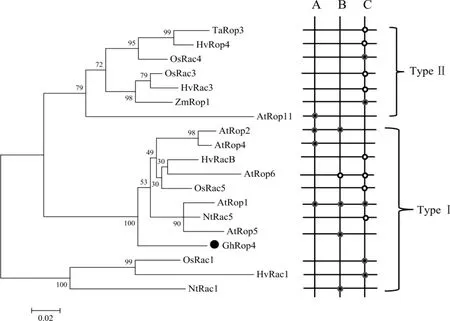
图3 Rop 蛋白聚类分析及生物学功能分析Fig.3 Cluster analysis and function analysis of Rop proteins
2.3 GhRop4基因表达特征分析
高盐(200 mol·L-1NaCl)、干旱、低温和黄萎病菌逆境胁迫处理均影响GhRop4 基因的表达,但是不同胁迫处理下基因表达模式不同。 在高盐处理下,GhRop4 基因在处理后3 h 的表达量最大, 处理后 1 h、6 h 和 12 h 的表达量均低于 0 h,其中在6 h 和12 h 的表达量较低(图 4A)。 干旱处理下,GhRop4 基因的表达量呈字母M 型趋势,在1 h 表达量升高,3 h 表达量降低,在 6 h 表达量急剧升高达到最大,随后又降低(图4B)。 低温处理后,随着胁迫处理时间的延长,GhRop4 基因表达量呈现逐渐下降趋势, 在3 h 时表达量最低,在6 h 表达量又急剧增加,12 h 表达量又有所下降(图4C)。 黄萎病菌侵染棉花后4 hGhRop4基因的表达量与0 h 表达量基本一致, 无显著差异; 在侵染后8 hGhRop4 基因的表达量显著增加,达到最大;处理后12 h 表达急剧降到最低;在24 h、48 h 基因表达量略微增加, 但仍明显低于0 h(图4D)。GhRop4 基因在不同组织中的表达分析结果表明,GhRop4 基因在检测的根、茎、叶、子叶和下胚轴中均有表达,在子叶中表达最高,其次是茎、叶、根,在下胚轴中的表达量最低(图4E),说明该基因的表达存在一定的组织特异性。

图4 GhRop4 基因表达谱分析Fig.4 Expression pattern analysis of GhRop4
3 讨论
棉花具有重要的经济和社会意义,对棉花的研究报道相对较多, 特别是在逆境胁迫反应方面。 如棉花GhWRKY91 基因介导转基因拟南芥叶片衰老及对干旱胁迫的响应[29]。 脱水素基因Gh_A05G1554 (GhDHN_03) 和 Gh_D05G1729(GhDHN_04) 介导植物耐受渗透胁迫和盐胁迫影 响 的 能 力[30]。 棉 花GhHB12[31]、GhLAC15[32]、GbaNA1[33]基因参与棉花抗黄萎病调控目前克隆到参与棉花抗逆的相关基因及信号传递途径主要涉及类受体蛋白、蛋白激酶、激素信号途径、转录因子调控、生化代谢酶、防卫基因合成等。 信号转导在植物生长发育和环境适应过程中扮演关键的调控作用。 在已报道的真核生物中,GTP 结合蛋白家族的Rho 家族蛋白是1 类信号转导的开关[34]。 目前棉花有6 个Rop基因的功能被报道, 其中 5 个基因GhRacA、GhRacB、GhRac9、GhRac13、GhRac1 功能研究集中在棉花纤维发育上[23-25],1 个GhRop6 基因参与了棉花对黄萎病抗性的调控[26]。 本研究利用生物信息学、PCR 技术从陆地棉中克隆到 1 个Rop基因, 命名为GhRop4。本研究克隆的GhRop4 基因与上述6 个进行序列比对发现, 相似性70%~88%, 表明GhRop4 基因是新的未报道的Rop基因。 DNAMAN 多重序列比对结果表明, 和已报道的Rop蛋白一样,含有 5 个(Ⅰ-Ⅴ)效应结构区,2 个开关区域,1 个Rop 插入序列及C 末端高度变异区(HVR),符合植物 Rop 蛋白的特征。
聚类分析中的7 个Ⅱ型Rop 蛋白除了At-Rop11 参与生长发育调控,其余6 个Rop 蛋白正调控或负调控植物抗病反应。Ⅰ型Rop 蛋白参与调控植物生长发育、抗逆反应和抗病性;聚类分析结果表明,GhRop4 蛋白属于Ⅰ型Rop 蛋白。与棉花GhRop4 蛋白聚类在同一亚组的AtRop1 和AtRop5 正调控植物的抗逆性[35],AtRop6 在这一过程发挥负调控作用[12]。 HvRacB、OsRac1、Os-Rac5 和NtRac5 参与植物对病原菌的入侵响应过程[19-20,22]。一般认为蛋白质具有相似的结构域可能具有相似的生物学功能,GhRop4 蛋白与上述蛋白聚为一类, 与它们的相似性均在80%以上,与OsRac5 和NtRac5 蛋白的相似性高达88%和90%, 推测GhRop4 蛋白可能在棉花抗逆性和抗病性反应中具有相似的功能。 因此本研究分析了GhRop4 基因在高盐、干旱、低温处理和黄萎病菌侵染后基因的表达情况,结果表明GhRop4 基因在盐和低温处理下,基因整体上呈现先下调再上调后有下调的趋势, 干旱和黄萎病菌处理后,基因呈现先升高后下降的表达趋势。 在这4 种逆境胁迫处理中GhRop4 基因的表达受干旱调控的程度最强,其次是黄萎病菌。 推测GhRop4 基因更多地参与了棉花对干旱的适应反应过程,对黄萎病菌的响应也显示出该基因在棉花抗黄萎病反应中也发挥作用。 上述结果暗示在后续的研究中应该更多地关注GhRop4 基因在棉花抗旱和抗黄萎病中的功能。 组织特异性分析表明,GhRop4 基因在棉花幼苗根、茎、叶、子叶和下胚轴中均有表达,与GhRac13[24]、GhRac1[25]、GhRop6[26]基因在检测的所有组织中均有表达的结果一致,但是存在一定的组织特异性,GhRop4 在子叶中的表达量较高,其余3 个已报道的Rop基因主要在棉花纤维中表达, 以上结果表明棉花GhRop基因在不同的组织中广泛地发挥着重要的调控功能。 本研究为进一步深入研究GhRop4 基因提供了重要的功能线索, 也首次揭示GhRop基因在抗旱中的功能。
4 结论
本文以陆地棉cDNA 为模板克隆到1 个小GTP 结合蛋白, 其开放阅读框为 588 bp, 编码195 个氨基酸, 包含 5 个效应结构区、2 个开关域、1 个Rop 插入序列及C 末端高度变异区,属于Ⅰ型Rop 蛋白,命名为GhRop4。组织表达分析表明,GhRop4 基因在棉花幼苗根、茎、叶、子叶和下胚轴中均有表达, 且在子叶中表达水平较高。同时,GhRop4 基因响应高盐、干旱、低温和棉花黄萎病菌等逆境胁迫。 本试验结果对开展棉花耐逆机制研究有重要意义。
猜你喜欢
环球时报(2022-09-20)2022-09-20
湖北农业科学(2022年11期)2022-07-18
作物学报(2022年9期)2022-07-18
农业科技与信息(2021年4期)2021-12-05
新农村(浙江)(2021年2期)2021-11-30
今日农业(2020年24期)2020-12-15
实用肿瘤学杂志(2020年4期)2020-12-08
农家科技下旬刊(2016年6期)2016-05-14
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
