
鄱阳湖丰水期水位波动对浮游动物群落演替的影响
2020-04-16 05:40胡旭仁欧阳珊秦海明
生态学报 2020年4期
吕 乾,胡旭仁,聂 雪, 欧阳珊,王 超,秦海明,2,3,*
1 南昌大学生命科学研究院流域生态学研究所和生命科学学院, 南昌 330031 2 曲阜师范大学生命科学学院, 曲阜 273165 3 南昌大学江西省流域生态演变与生物多样性重点实验室, 南昌 330031 4 中国水产科学研究院珠江水产研究所, 广州 510380
浮游动物在湖泊生态系统食物网的物质循环和能量传递的过程中发挥着重要作用,对于淡水生态系统的稳定性具有重要意义[1- 2],揭示湖泊系统中的浮游动物群落结构特征有助于认识湖泊生态系统中物质和能量的流动[3]。关于环境因子对浮游动物群落结构具有影响已经被很多学者报道,叶绿素a含量、化学需氧量、水温、总磷、总氮、水体的酸碱度和电导率是影响浮游动物群落结构的主要水环境因子[4- 6]。国际上的一些研究发现水文特征可通过影响生境条件而对浮游动物的群落多样性产生影响[7- 8],水位变化对浮游动物群落结构及其动态变化具有显著影响[9]。而在鄱阳湖进行的浮游动物研究发现水文变化是浮游甲壳类群落结构变化的重要因素之一[10]。
鄱阳湖是长江流域最大的通江湖泊,年内水位变幅很大,季节性的洪水涨退和周期性的湖水更换致使形成了典型的洲滩湿地。此外,鄱阳湖生物多样性资源十分丰富,而且具有悠久的人类活动历史,诸多因素使得鄱阳湖成为一个人地交互作用显著的动态湖泊[11- 12]。随着人类对环境的影响加剧,鄱阳湖生态系统正面临着严峻考验并发生了一系列变化,其中的生物组成、群落结构均随之发生着改变[13]。了解浮游动物和鱼类等水生生物的丰度及其空间分布状况对理解湖泊生态系统功能至关重要[9]。因此对当前鄱阳湖中浮游动物的种类组成、时空分布和群落结构特征进行研究非常有必要。本研究的主要目的是:1) 研究丰水期鄱阳湖的水位剧烈波动是否对浮游动物的群落产生直接影响?2) 调查丰水期水位波动过程中浮游动物群落的演替特征如何?3) 分析对浮游动物群落演替具有重要影响的环境因子有哪些?通过对保护区子湖中浮游动物群落结构演替特征的研究及水位变化对浮游动物群落影响的分析,可以为保护区制定合理的管理措施,保护鄱阳湖生态系统完整性和稳定性供科学依据。
1 材料方法
1.1 采样地点
鄱阳湖 (28° 24′—29° 46′N, 115° 49′—116° 46′) 位于长江下游,属于典型的季节性吞吐湖泊,受亚热带季风性气候、长江和五河 (赣江、修河、抚河、饶河、信江) 河水注入的影响,水位年变幅大。每年4月至9月为丰水期,10月至次年3月为枯水期,丰水和枯水季节湖水位变化最大可达9—14 m[14]。江西鄱阳湖国家级自然保护区位于鄱阳湖西北部,修河和赣江两河交汇处,保护区辖区内9个子湖泊 (图1),年平均降雨量1400—1900 mm,年平均气温16.7—17.7 ℃。

图1 江西鄱阳湖国家级保护区地理位置及沙湖浮游动物采样点设置Fig.1 Location of Jiangxi Poyang Lake National Nature Reserve and the zooplankton sampling points in Shahu Lake

图2 2012年6月1日至8月31日沙湖的水位变化及采样当天水位Fig.2 Diurnal variation of water level from June 1 to August 31, 2012 and water level on the sampling days in Shahu Lake
采样地点沙湖 (115° 56′ E, 29° 10′ N) 是江西鄱阳湖国家级自然保护区内的一个核心子湖泊,属于典型的浅水碟型子湖,面积约为14 km2。枯水季节与修河和主湖断开,形成独立的子湖泊,丰水季节通过修河与主湖连成一个大鄱阳湖,丰水期的最高水位可达20.58 m[15]。
1.2 样本采集
在2012年夏季丰水期鄱阳湖水位剧烈波动上涨期间 (6月1日至8月31日),分别于6月24日、7月7日和8月27日在鄱阳湖的1个子湖沙湖进行浮游动物定量采集,三次采样均处于水位下降末期,水位非常接近 (17.64 m、17.60 m、17.68 m,图2) 且子湖与主湖保持连通。根据鄱阳湖国家级自然保护区管理局的长期水位监测结果,当水位高于17.3 m时,沙湖通过修河与鄱阳湖主湖区连通。在沙湖敞水区的不同水深区域分别设置4个采样点 (图1),每个采样点间隔20 m设置3个采样重复,仅采集水表层下50 cm处的水样,采样方法参照《淡水浮游生物研究方法》[16]。
1.3 样品处理
实验室内使用虎红钠盐将浮游动物样品染色,采用体积法抽样[7],解剖镜下鉴定计数,剩余样品用4%福尔马林溶液固定保存。物种分类和鉴定参照《中国动物志 淡水桡足类》、《中国动物志 淡水枝角类》、《中国淡水轮虫志》和《中国淡水生物图谱》[17-20]。其中桡足类无节幼体未鉴定到物种水平,分析时作为一个分类群统计。
1.4 数据分析
物种优势度计算公式如下:
Y=ni×fi/N
其中,Y表示物种优势度,ni为第i种的个体数,N为所有个体总数的和,fi为该种出现的频度,Y≥0.02定义为优势种[21]。
通过辛普森指数、物种丰富度指数、物种均匀度指数分析3个月份浮游动物多样性指数的变化。多样性指数计算公式如下:
Shannon-Wiener 指数:
H′= -∑Pi×lnP
Margalef 指数:
D=(S-1)/lnN
Pielou 指数:
J=H′/lnS
上式中Pi=ni/N,ni为i种的个体数,N为所有种类总个体数,S为物种数[5]。
使用ArcGIS 10.0绘制夏季沙湖水深分布图。在STATISTICA 6.0软件中,运用单因子方差分析检验不同月份之间的水体理化因子、浮游动物密度、种类数差异,采用最小显著差异法 (LSD) 进行多重比较,P< 0.05为差异显著。使用软件Primer 5.0进行浮游动物群落多维无度量分析 (NMDS) 和群落相似性分析 (ANOSIM),并基于多度使用 Bray-Curtis 相似性系数对浮游动物群落进行聚类。在MATLAB R2014a软件中,使用Kohonen自组织映射神经网络图(SOM)对浮游动物群落进行聚类[22]。使用软件Canoco for Windows 4.5对浮游动物群落和环境因子进行典型相关分析 (CCA)。所有数据在统计分析前进行了log (x+1)转换。
2 研究结果
2.1 理化性质
7月份的水温 (30.49 ℃) 显著高于6月份 (27.60 ℃) 和8月份 (28.15 ℃)。8月份水体溶解氧 (8.11 mg/L) 略高于7月份 (7.66 mg/L),显著高于6月份 (7.04 mg/L)。6月份水体pH值 (7.24) 显著低于7月份 (7.51) 和8月份 (7.43)。6月份水体叶绿素a含量 (12.78 μg/L) 显著高于7月份 (8.55 μg/L) 和8月份 (10.28 μg/L)。8月份水体的电导率 (1194 μs/cm) 和浊度 (9.48 NTU) 显著高于6月份 (电导率99 μs/cm;浊度6.22 NTU) 和7月份 (电导率83 μs/cm;浊度2.71 NTU) (图3)。

图3 沙湖2012年夏季3个月份的水理化因子Fig.3 Physicochemical parameters of Shahu Lake in the three months of summer, 2012
2.2 浮游动物种类组成
2012年夏季3个月份在沙湖的3次调查共采集到浮游动物65种,隶属于2个动物门,20科。其中轮虫52种、桡足类7种、枝角类6种。轮虫为绝对优势类群,占总数量的80.00% (表1),在3个月中的物种数均超过70.00%,枝角类物种数变化不大,桡足类物种数逐月增加。
2.3 浮游动物优势物种组成变化
计算浮游动物的优势度,发现优势度指数大于等于0.02的优势物种有18种,其中轮虫15种,枝角类1种,桡足类2种 (表2)。3个月的优势种差异较大,6月份有9种优势物种,除桡足类无节幼体,外其他均为轮虫。7月份有10种优势物种,除1种枝角类和桡足类无节幼体外其他均为轮虫。8月份有8种优势物种,其中轮虫5种,枝角类和桡足类各2种。3个月的共同优势种仅有1种,为盖氏晶囊轮虫Asplanchnagirodi。优势物种广布多肢轮虫Polyarthravulgaris、长肢多肢轮虫Polyarthradolichoptera和螺形龟甲轮虫Keratellacochlearis仅在6月份和7月份出现,圆筒异尾轮虫Trichocercacylindrica和长额象鼻溞Bosminalongirostris仅在7月份和8月份出现。

表1 沙湖2012年夏季3个月份浮游动物种类及百分比
2.4 浮游动物的密度与生物量
浮游动物的总密度在7月份最高 (995.06±94.76 个/L),显著高于6月份(302.25±53.32 个/L) 和8月份 (203.49±24.03 个/L)。轮虫的密度在7月份显著高于6月份和8月份,枝角类和桡足类均在8月份出现最高密度,显著高于7月份和6月份的密度 (P<0.05) (图4)。
浮游动物的总生物量在7月份 (1.22±0.11 mg/L) 最高,8月份 (0.99±0.11 mg/L) 次之,6月份 (0.56±0.04 mg/L) 最低。轮虫在7月份的生物量显著高于6月份和8月份,而枝角类和桡足类的生物量均在8月份最高,6月份最低 (P<0.05) (表3)。

表2 沙湖2012年夏季3个月份浮游动物优势物种及优势度指数
“-”表示未发现该种类

图4 沙湖2012年夏季3个月份的浮游动物密度 (n=36)Fig.4 Zooplankton density of Shahu Lake in the three months of summer, 2012

表3 沙湖2012年夏季3个月份的浮游动物生物量
2.5 浮游动物的群落结构
基于浮游动物多度的聚类分析和自组织映射神经网络图均揭示夏季3个月份浮游动物区分为明显的3个群落:6月群落、7月群落和8月群落 (图5)。相似性分析 (ANOSIM) 进一步揭示3个月份浮游动物群落之间具有显著差异 (R=0.983,P=0.001)。
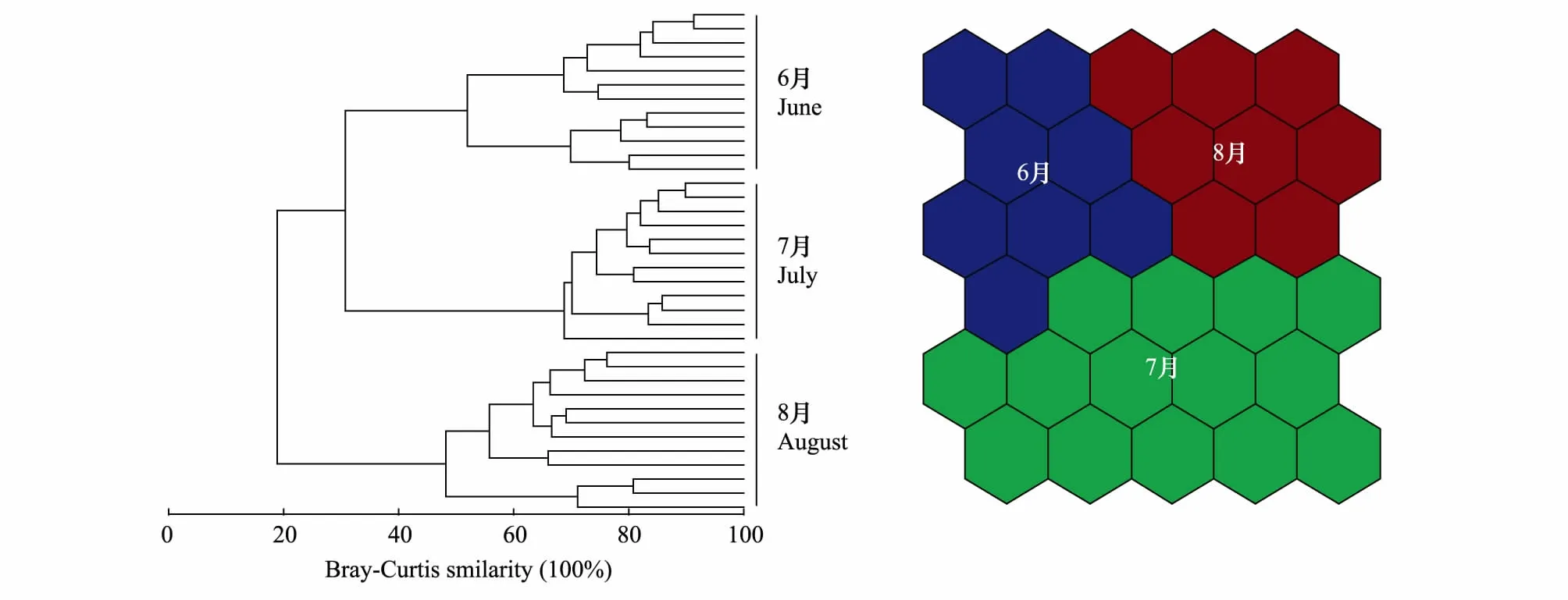
图5 基于多度数据的6月、7月和8月浮游动物群落聚类和自组织映射神经网络图Fig.5 Clustering of zooplankton communities by Bray-Curtis similarity measure and self-organizing maps in Shahu Lake on April, July and August based on abundance data
2.6 浮游动物群落结构与环境因子的关系
典型对应分析 (CCA) 结果显示,水温、溶氧、pH值、电导率、叶绿素a含量和浊度6个水体理化因子对浮游动物群落变化的总方差贡献率为85.6%,第一排序轴的特征值为0.285,方差贡献率为39.8%;第二排序轴特征值为0.214,方差贡献率为29.9%。蒙特卡罗检验发现水温、电导率、浊度和溶氧与浮游动物群落结构变化显著相关 (水温:F=5.24,P=0.002;电导率:F=6.62,P=0.002;浊度:F=1.41,P=0.03;溶氧:F=1.53,P=0.02)。3个月份的浮游动物明显的各自聚为一个群落,6月份浮游动物群落分布在第II象限,与叶绿素a含量呈显著正相关关系,7月份浮游动物群落分布在第III象限,与水温呈显著正相关关系,8月份浮游动物群落分布在第I和第IV象限,与水深、电导率、浊度和溶氧呈正相关关系 (P<0.05, 图6)。4种优势物种分布在第I象限,5种分布在第II象限,另4种分布在第III象限,还有3种分布在第IV象限。蒙特卡洛检验结果表明,盖氏精囊轮虫与叶绿素a含量呈正相关关系 (P<0.05),与水温、PH和溶氧负相关 (P<0.05),与电导率、水体浊度和溶解氧的无明显相关性关系。

图6 浮游动物种类、采样点和水体理化性质典型对应分析,空白三角形 (△) 表示浮游动物的种类,黑色三角形 (▲) 表示优势物种Fig.6 Canonical correspondence analysis tri-plot of zooplankton species/taxon, sampling stations and environment conditions. Blank triangles (△) indicate zooplankton species. Black triangles (▲) indicate dominant species
3 讨论
3.1 水位波动过程中环境因子的变化
已有的研究显示温度与水体pH的关系的研究结果显示温度与pH值呈负相关关系[23],本研究未发现相似的规律。另有研究指出天然水体pH值的变化受降水和河流含沙量的影响明显[24],本研究在夏季丰水期开展,由于鄱阳湖水位的剧烈波动,水体中的含沙量存在明显变化,对水体的pH值影响较大。另有研究报道叶绿素a的浓度与总磷浓度和水温呈正相关,与透明度呈负相关[25]。水位波动对水体叶绿素a浓度具有显著的影响[26],鄱阳湖水位的波动势必也会对水体中的叶绿素a浓度产生影响。另有研究表明溶解性氮浓度、水体透明度和总氮浓度与鄱阳湖的水位变化显著相关[27]。本研究发现叶绿素a浓度与浊度呈正相关关系,这与吕唤春等的研究结果[25]一致,而叶绿素a浓度与水温呈负相关关系则与前期的研究结果不同。
通常情况下水体的溶解氧与温度呈负相关关系[28],但是本研究的结果不完全符合这个规律。在6月份采样时,沙湖已经与鄱阳湖主湖连通接近20天,大面积的洲滩湿地植物被淹没后腐烂发酵会大量消耗水体中的溶解氧,这造成了6月份采样时水体的溶解氧处于较低水平。夏季沙湖一直保持较高的水位,洲滩上的湿地植物一直处于被淹没状态,随着湿地植物的不断腐烂发酵,水体溶解氧的消耗逐渐减少。此外,根据水位监测数据可以发现,6月至8月沙湖的水位一直处于剧烈波动上升或下降,水体的扰动也增加了溶解氧的含量。可以推测这是造成沙湖的水体溶解氧在夏季3个月份逐渐升高的主要原因。
3.2 浮游动物种类组成变化
夏季沙湖浮游动物的物种数月变化与其他湖泊的变化并不一致,沙湖中浮游动物的物种数呈先减少再增加的变化特征,而太湖浮游动物的物种数在夏季3个月基本相同[29]。本研究对沙湖浮游动物调查结果表明轮虫为优势类群,与历史调查研究结果保持一致,3个月的轮虫物种数的比例均超过70%。有关鄱阳湖的浮游动物研究结果均表明轮虫是优势类群[30-32]。已有的研究显示浮游动物第一优势物种的优势地位特别明显,例如在南海北部大亚湾的研究指出浮游动物的优势物种季节变化明显,优势物种的变化主要受其生活环境影响,且第一优势物种的优势地位特别明显[33];对杭州西湖浮游动物群落变化的研究中也发现第一优势物种的优势地位特别明显[34]。本研究的结果与上述报道一致,夏季3个月份浮游动物的优势物种变化明显,在3个月均为优势种的仅有1种,且各月份第一优势物种的密度显著高于第二优势物种。相关研究对2012—2016年沙湖的浮游动物优势物种进行分析发现,沙湖浮游动物的优势物种在年际间存在明显差异[30],可见沙湖浮游动物优势物种在年内和年际间的差异都很明显。
中国动物志中有记述,长额象鼻溞从6月开始种群逐渐消失,直到九月才再次出现[19],而本研发现在7月和8月长额象鼻溞均为优势物种,密度可达261.33 个/L和372.73 个/L,可能的原因是夏季鄱阳湖剧烈的水位波动引起了水环境的巨大变化,从而导致长额象鼻溞的繁殖周期发生了一定改变。环境因子对浮游动物生殖周期的影响早已有研究确认,例如环境因子变化会影响枝角类的生殖周期[35],且对萼花臂尾轮虫的繁殖也具有显著影响[36],因此,水位的剧烈波动可能通过对水环境因子的影响间接影响浮游动物的繁殖周期,从而导致浮游动物的群落结构产生差异。
3.3 浮游动物群落演替的影响因素
浮游动物对水环境变化十分敏感,环境改变会促使浮游动物群落结构发生演替[37]。本研究通过对环境因子与浮游生物群落的典型相关分析 (CCA) 发现由于环境因子的差异性,导致3个月的浮游动物群落呈现显著差异。相关研究发现水温对浮游动物群落变化具有重要影响[32],本研究发现7月份浮游动物的密度与水温呈显著正相关,而在6月份和8月份浮游动物的密度与水温呈负相关关系。有关浮游动物与水环境因子关系的研究发现电导率和溶氧是影响浮游动物的主要环境因子[38],该研究的结果支持这个结论。本研究在夏季鄱阳湖水位变化时期开展,水位的剧烈波动导致湖泊底质再悬浮[39],引起水体浊度增加,而浊度与浮游动物的密度具有显著相关关系[20],浊度的改变直接影响了浮游植物和沉水植物的生长[40],进而对以浮游植物为食的次级消费者浮游动物产生影响。
水位变动是影响湖泊生态系统状态变化的主要因素[41]。国际上有关研究指出水位变动对浮游动物的多样性、分布和动态都具有明显的影响[42, 43]。另有研究发现不同水位时期浮游动物的群落结构具有明显差异,中等水位时期浮游动物的种类、丰富度和多样性显著高于高水位和低水位时期[44]。本研究发现轮虫对浮游动物总密度的贡献率最高,是由于水位波动过程中湖泊水力滞留时间短,轮虫繁殖速度快,生命周期短,可以很快的适应快速变换的水生态环境,形成优势类群[44, 45]。此外,高水位时期由于沙湖与主湖的连通,使得浮游食性的鱼类大量进入子湖觅食,对浮游动物的密度产生直接影响。在鱼类的捕食压力下,体型较大的枝角类和桡足类被捕食的概率更高,因此个体较小的轮虫具有相对较高的生存优势[46]。
依据上述讨论,很难确定水位变化是否对夏季浮游动物的群落演替产生直接影响。查阅文献发现,在太湖和红枫湖开展的周年浮游动物群落动态研究结果表明:6月至8月浮游动物群落的演替特征表现为密度逐渐下降 (6月份>7月份>8月份);生物量先降低再增加 (6月份>8月份>7月份)[29, 47]。而本研究发现夏季3个月份鄱阳湖1个子湖中浮游动物的群落演替特征表现为密度先升高后降低 (7月份>6月份>8月份,图4);生物量先升高后降低 (7月份>8月份>6月份,表3),特别是沙湖中枝角类和桡足类的密度变化呈现逐月上升的变化特征,这与太湖的调查结果完全不同。太湖和红枫湖是相对封闭的内陆湖泊,在夏季与连通性的沙湖具有完全不同的水文特征。基于沙湖的浮游动物群落演替与已报道的太湖和红枫湖完全不同,我们认为丰水期鄱阳湖的水位波动是影响沙湖浮游动物群落演替的一个直接而重要的因素。
4 结论
夏季水位波动过程中鄱阳湖子湖内的浮游动物群落结构在时间上 (月份之间) 发生明显演替,呈现轮虫密度逐渐降低,枝角类和桡足类密度逐渐增加的变化特征。水位的剧烈波动导致水环境发生快速变化,对一些浮游动物的生长繁殖活动产生影响,从而导致了浮游动物群落的快速演替。
猜你喜欢
水利水电快报(2022年7期)2022-07-18
星星·散文诗(2020年10期)2020-12-03
科学养鱼(2020年10期)2020-11-23
课外语文·中(2020年2期)2020-03-24
课外语文(2020年5期)2020-03-14
现代出版(2019年6期)2020-01-14
知识窗(2019年4期)2019-04-26
老友(2017年4期)2017-02-09
林业与生态(2016年2期)2016-02-27
军事历史(1985年4期)1985-08-20
