
不同生境中刺萼龙葵空间点格局分析
2020-04-14 04:59陈菁马方舟张彦静王晨彬徐海根
南方农业学报 2020年2期
关键词:生境
陈菁 马方舟 张彦静 王晨彬 徐海根

摘要:【目的】掌握刺萼龍葵种群的分布格局,为刺萼龙葵的有效控制提供理论依据。【方法】采用点格局方法,运用Ripleys L函数对不同生境(河滩、草原和荒地)中的刺萼龙葵种群进行点格局及不同植株类型间的关联性分析。【结果】3种生境中的刺萼龙葵不同植株大小类型数量排序均为大型植株>中型植株>小型植株,大型和中型植株以河滩生境最多,草原生境最少。3种生境中,刺萼龙葵在小尺度上均表现为聚集分布,在大尺度上表现为随机分布或均匀分布。在河滩生境中,仅中型植株表现出微弱的聚集性;而在草原生境和荒地生境中,在中小尺度上大型植株和中型植株均表现出聚集性,且大型植株相较于中型植株表现出更强的聚集性。在河滩生境中,仅中型植株与小型植株间在小尺度上表现出显著正相关性,大、中、小3种植株类型间随着尺度的增大,不断趋于无相关性或显著负相关;在草原生境和荒地生境的中小尺度内,大、中、小3种刺萼龙葵植株类型间均表现出一定的显著正相关性,随着尺度的增大,不断趋于无相关性或显著负相关。【结论】入侵植物刺萼龙葵的群落空间格局在不同资源环境条件下均呈现有利于种群扩散的分布特征,且不同株型间易形成互利的生态关系,是其成功入侵的主要驱动因素之一。
关键词: 刺萼龙葵;生境;点格局分析;植株大小类型;空间关联性
中图分类号: S451.1 文献标志码: A 文章编号:2095-1191(2020)02-0342-08
Spatial point pattern analysis of Solanum rostratum
Dunal in different habitats
CHEN Jing1,2, MA Fang-zhou2*, ZHANG Yan-jing2, WANG Chen-bin2, XU Hai-gen1,2*
(1College of Biology and Environment, Nanjing Forestry University, Nanjing 210037, China; 2Nanjing Institute of Environmental Sciences, Ministry of Ecology and Environment / National Key Laboratory of Environmental
Protection and Biosafety, Nanjing 210042, China)
Abstract:【Objective】In this study,the effects of spatial distribution pattern of Solanum rostratum Dunal population on the invasion process was taken as the focus of attention. The aim of this study was to providea better understanding of the theoretical basis for S. rostratum control. 【Method】Point pattern analysis(PPA) was adoptedto examine spatial point pattern and the correlation among populations of S. rostratum in different habitats(beach,grassland and wasteland) using Ripleys L function. 【Result】For the three habitats, the number of S. rostratum seedlings in different types ranked as large plants>medium plants>small plants. The numbers of large and medium plants in beach habitat were the largest, and in grassland habitat was the smallest. Populations of S. rostratum in three habitats showed aggregation distribution on small scale,and random or uniform distribution on large scale. In beach habitats,only medium plants of S. rostratum showed slight aggregation,while large and medium plants showed aggregation on small scalein grassland and wasteland habitats,besides,the large plants showed stronger aggregation than the medium plants. In beach habitats,only the medium plants and the small plants showed positive correlation in the small scale range,and the large,medium and small plant types ten-ded to show no correlation or negative correlation with the increase in the scale range. In grassland and wasteland habitats,there was a positive correlation among the three plant types of S. rostratum in the medium and small scale range,however,no correlation or negative correlation as the increase of the scale range. 【Conclusion】The spatial pattern of invasive plant S. rostratum community in different resource and environment conditions show distribution characteristics conducive to population diffusion,and the mutually beneficial ecological relationship between different plant types is formed,these are the main driving factors for its successful invasion.
Key words: Solanum rostratum Dunal; habitat; point pattern analysis; plant size; spatial correlation
Foundation item:National Key Research Project of China(2017YFC1200100);Basic Research Project of Central Public Welfare Research Institutes(GYZX180203)
0 引言
【研究意义】刺萼龙葵(Solanum rostratum Dunal)又名黄花刺茄,隶属于管花目茄科茄属,是一种入侵性极强的一年生恶性杂草。该入侵植物原产于北美洲,通过混杂在进口粮食中传入,1895年在香港首次被发现,现已扩散至欧洲、亚洲、非洲和大洋洲的近20个国家和地區。1981年,刺萼龙葵在我国大陆地区首次发现于辽宁省朝阳县,现已逸生于吉林、河北、山西、北京、新疆和内蒙古等省(区、市),且有逐渐向西、向南扩张的趋势,其潜在分布区极为广泛,覆盖全国大部分地区,尤其以华北、华东、华中、东北和西南地区为高风险分布区(关广清,2000;钟艮平等,2009;Horvitz et al.,2017)。刺萼龙葵的繁殖系统在原产地以异交为主,而在我国北京和张家口入侵地区可自发自交,繁殖能力极强(张丽娟和娄安如,2018),其植株具尖刺,植株含龙葵碱,可导致人畜中毒。鉴于刺萼龙葵较强的入侵性、扩散性及对生态环境和农林业生产的危害性,我国已将其定性为检疫杂草,并列入《中华人民共和国进境植物检疫性有害生物名录》和环境保护部公布的《中国自然生态系统外来入侵物种名单(第四批)》,以加强对其检疫和防控监管。因此,分析刺萼龙葵种群空间分布格局,收集不同生境下刺萼龙葵入侵和扩张趋势及相关种群生物学信息,对掌握其暴发规律,揭示入侵机制及进行有效防控管理均具有重要意义。【前人研究进展】植物种群的空间分布格局是指种群个体在空间的分布情况,反映种群个体间在空间水平的相互关系(张金屯,1998)。种群空间分布格局及其关联性对种群自身的生长、繁殖和资源利用等有显著影响,也是种群自身生物学特性对其生长环境条件适应和选择的结果(Druckenbrod et al.,2005;王梅等,2015;李国春等,2017)。一般来说,植物种群在群落中的分布有3种类型,即随机分布、聚集分布和均匀分布。空间分布格局的研究方法包括样方法、距离法和点格局分析法等(李明辉等,2003)。目前关于刺萼龙葵空间分布规律的研究多是基于较大空间尺度范围,钟艮平等(2009)基于GARP生态位模型对刺萼龙葵在我国的潜在分布区进行了预测;王瑞等(2018)基于Maxent模型分析了刺萼龙葵在我国的地理分布格局,并划定了早期监测预警区域。而在较小尺度范围内,如单个种群内,关于刺萼龙葵个体间的分布规律研究较少。生态学过程具有尺度依赖性,若不考虑空间尺度因素,就无法揭示这些生态学过程的相对重要性(Wiegand and Moloney,2013),因此,格局与尺度有密切关系。传统研究方法只能分析单一尺度空间分布格局,种群分布类型与空间尺度的关系无法全面反映出来(杨云方等,2013;胡砚秋等,2016),点格局分析法克服了传统方法的不足,得到快速发展并应用于格局分析中,成为空间格局分析最主要的方法(杨洪晓等,2006)。点格局分析方法是由Ripley于1977年首先提出,后经Diggle等完善发展而逐步形成的一种分析方法(Ripley,1976,1977)。近年来,点格局分析方法已广泛用于生态学和植物学等领域(时培建等,2010;Wilschut et al.,2015;樊登星和余新晓,2016;李国春等,2017),但目前还未发现有入侵学领域的相关研究。基于点格局的刺萼龙葵种群研究可揭示其分布规律(聚集分布、随机分布或均匀分布),有研究表明,刺萼龙葵的成功入侵与其种群密度有关,在入侵过程中存在植株间的正向密度依赖效应,即聚集性分布有利于生物的成功入侵(张丽娟和娄安如,2018)。【本研究切入点】目前关于刺萼龙葵种群个体间分布规律的研究较少,本研究针对入侵早期刺萼龙葵种群密度相对较小的特点,在小尺度范围内开展轻中度暴发阶段的刺萼龙葵种群点格局研究,分析不同生境条件下入侵植物种群的空间分布适应性策略和不同株型间的生态关系。【拟解决的关键问题】基于点格局分析理论,收集不同生境中刺萼龙葵入侵和扩张趋势及相关种群生物学信息,有助于分析其暴发规律,从刺萼龙葵空间分布格局对入侵过程影响的角度揭示其入侵机制,为刺萼龙葵的有效防控管理提供基础资料和科学依据。
1 材料与方法
1. 1 样地调查
试验地(东经122°15′4″,北纬46°0′25″)位于内蒙古东北部的乌兰浩特市,属中温带半干旱大陆性季风气候区,海拔180 m,年平均气温5.5 ℃,年平均降水量372.4 mm。在乌兰浩特市及其周边共设3个刺萼龙葵调查样地,分别代表3种典型生境(河滩、草原和荒地)。在样地内选择地势平坦、群落外貌均一且具有代表性的5 m×5 m刺萼龙葵样方,并将其进一步分割成100个50 cm×50 cm的小样方,采用相机对小样方进行垂直拍摄,保证小样方的4个角点恰好能进入取景器。拍摄完毕后,将采集的300张照片按一定顺序进行编号,便于进行后续处理。
1. 2 植株类型划分
现场调查时,以刺萼龙葵植株高度作为表征植株大小类型的指标进行测量统计,按株高将刺萼龙葵划分为3个等级:株高>60 cm为大型植株,60 cm≥株高>30 cm为中型植株,株高≤30 cm为小型植株。
1. 3 点格局分析方法
1. 3. 1 种群分布格局 Ripleys L函数可分析不同生境中刺萼龙葵种群的空间分布格局。Ripleys L函数由RipleyK函数转化而来,RipleyK函数的计算公式(Diggle,2003;Wilschut et al.,2015)如下:
K(r)=[An2][i=1nj=1j≠1nwij-1Ir(uij)]
式中,A为样方面积;n为个体数;r为距离尺度;uij为i和j两点间的距离;Ir(uij)为指示函数,当uij≤r时,Ir(uij)=1,当uij>r时,Ir(uij)= 0;wij为权重,用于进行边缘校正。
为更直观地解析种群空间格局,可用Ripleys L函数代替RipleyK函数:
L(r)=[K(r)/π-r]
当L(r)=0,种群呈随机分布;当L(r)>0,说明在以任意点为圆心、半径为r的圆内个体数比在空间完全随机情况下多,种群呈聚集分布;当L(r)<0,说明在以任意点为圆心、半径为r的圆内个体数比在空间完全随机情况下少,种群呈均匀分布。采用Monte Carlo模拟99%的置信区间进行显著性检验:L(r)位于上包迹线以上,表示种群呈聚集分布;L(r)位于下包迹线以下,表示种群呈均匀分布;L(r)位于上下包迹线之间,表示种群呈随机分布。
1. 3. 2 空间关联性关系 空间关联性关系可解析不同大小类型刺萼龙葵个体的空间关联性。空间关联性关系的公式(樊登星和余新晓,2016;梅军林等,2017)如下:
K12(r)=[An1n2][i=1n1j=1n2wij-1Ir(uij)]
式中,n1和n2分别表示类型1和类型2的个体数,i和j分别表示类型1和类型2的个体。用L12(r)进一步取代K12(r),其公式为:
L12(r)=[K12(r)/π-r]
当L12(r)=0,表明两种类型在r尺度下无关联性;当L12(r)>0,表明两种类型为空间正相关;当L12(r)<0,表明两种类型为空间负相关。采用Monte Carlo模拟99%的置信区间进行显著性检验,当L12(r)位于上包迹线以上时,二者呈显著正相关;当L12(r)位于下包迹线以下时,二者呈显著负相关;L12(r)位于上下包迹线之间时,二者相互独立。
1. 4 统计分析
Ripleys L函数的运算过程通过Programita软件完成,采用Excel 2013制图。
2 结果与分析
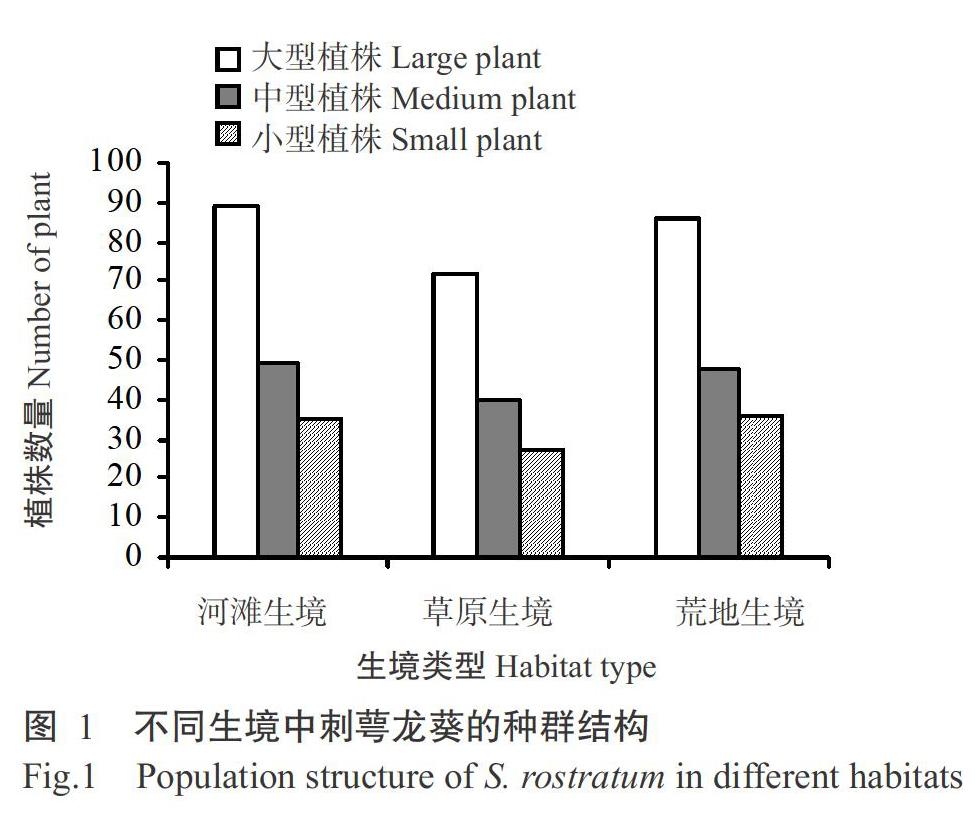
2. 1 不同生境中刺萼龙葵的种群结构
3种不同生境中刺萼龙葵个体数量远高于伴生植物,盖度约65%。3种生境中刺萼龙葵个体密度差别不明显,其中河滩生境中的个体密度最大,为6.92株/m2,其次是荒地生境,个体密度为6.80株/m2,草原生境中个体密度最小,为5.56株/m2。3种生境中刺萼龙葵不同植株大小类型数量排序均为大型植株>中型植株>小型植株,其中小型植株在3个生境中所占的比例约20%,中型植株所占比例约28%,大型植株所占比例最高,约51%。大型植株和中型植株均以河滩生境最多,草原生境中的大型植株和中型植株最少,荒地生境中的小型植株较多(图1)。
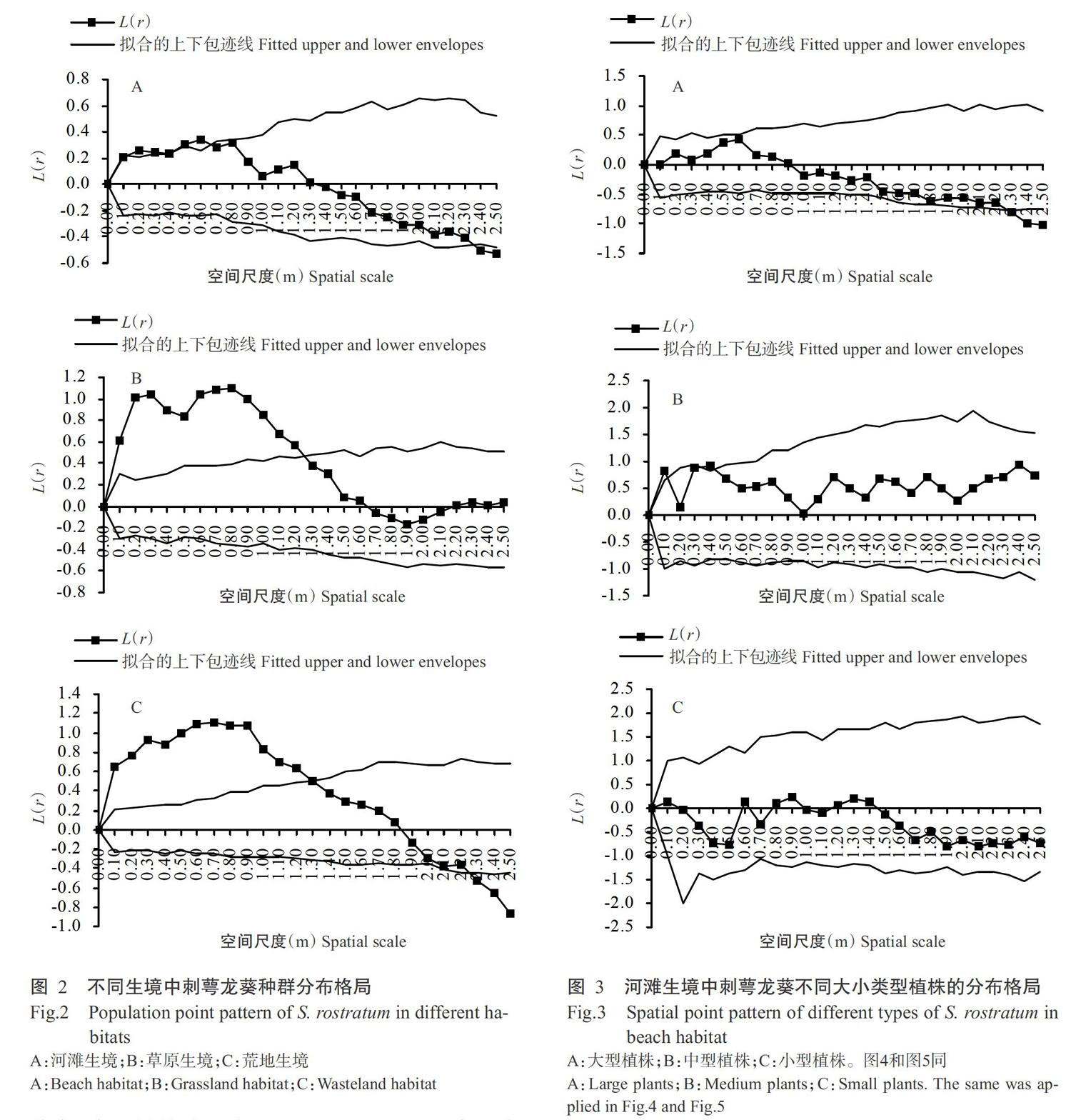
2. 2 不同生境中刺萼龙葵种群分布格局
不同生境中刺萼龙葵种群分布格局的分析结果表明,河滩生境中的刺萼龙葵在0.10~0.65 m尺度内表现为聚集分布,在2.35~2.50 m尺度内表现为均匀分布,在其他尺度上表现为随机分布(图2-A);草原生境中的刺萼龙葵在0~1.25 m尺度内表现为聚集分布,在1.25~2.50 m尺度内表现为随机分布(图2-B);荒地生境中的刺萼龙葵在0~1.30 m尺度内表现为聚集分布,在1.30~2.25 m尺度内表现为随机分布,在2.25~2.50 m尺度内表现为均匀分布(图2-C)。
2. 3 不同生境中刺萼龙葵不同植株大小类型种群分布格局
河滩生境中,刺萼龙葵大型植株在2.30~2.50 m尺度内表现为均匀分布,在其他尺度内表现为随机分布;中型植株除了在0~0.10 m及0.40 m尺度上表现为聚集分布外,在其他尺度内均表现为随机分布;小型植株在研究尺度范围内均表现为随机分布(图3)。草原生境中,刺萼龙葵大型植株在0~1.30 m尺度内表现为聚集分布,在1.30~2.50 m尺度内表现为随机分布;中型植株除了在0.40 m尺度上表现为聚集分布外,在其他尺度范围内均表现为随机分布;小型植株在研究尺度范围内均表现为随机分布(图4)。荒地生境中,刺萼龙葵大型植株在0~1.75 m尺度内表现为聚集分布,在1.75~2.50 m尺度内表现为随机分布;中型植株在0~0.70 m尺度内表现为聚集分布,在0.70~2.00 m尺度内表现为随机分布,在2.00~2.50 m尺度内表现为均匀分布;小型植株在0~0.40 m尺度内表现为聚集分布,其他尺度范围内表现为随机分布(图5)。
2. 4 不同生境中刺萼龙葵不同植株大小类型的空间关联性分析结果
刺萼龙葵种群在不同生境中不同植株大小类型间的空间关联性分析结果表明,在河滩生境中,研究尺度内的大型植株与中型植株在0~0.20 m尺度內呈显著正相关,在2.20~2.50 m尺度内呈显著负相关,其他尺度内无相关性;大型植株与小型植株在0~2.10 m尺度内无相关性,在2.10~2.50 m尺度内呈显著负相关;中型植株与小型植株在0.40 m及1.10~2.50 m尺度内无相关性,在其他尺度内呈显著正相关(图6)。在草原生境中,大型植株与中型植株在0.50 m和1.05~2.50 m尺度范围内无相关性,在其他尺度范围内呈显著正相关;大型植株与小型植株在0.25~0.95 m尺度范围内呈显著正相关,在其他尺度范围内无相关性;中型植株与小型植株在0.15~0.95 m尺度范围内呈显著正相关,在其他尺度内无相关性(图7)。在荒地生境中,大型植株与中型植株在0.15~1.05 m尺度内呈显著正相关,在2.40~2.50 m范围内呈显著负相关,在其他尺度范围内无相关性;大型植株与小型植株在0~0.35 m和0.50~0.95 m尺度范围内呈显著正相关,在2.10~2.50 m尺度范围内呈显著负相关,在其他尺度范围内无相关性;中型植株与小型植株在0~0.70 m尺度范围内呈显著正相关,在1.90~2.50 m尺度范围内呈显著负相关,在其他尺度范围内无相关性(图8)。
3 讨论
植物种群的空间分布格局不仅与其自身生物学特性(繁殖特性、竞争机制和资源利用率等)相关,还受环境因素,包括地形、土壤、水分和光照等异质性制约(时培建等,2010;赵欣胜等,2011;Li and Zhang,2015;Nguyen et al.,2016)。在3种生境内,随着研究尺度的增大,刺萼龙葵总体上呈现先聚集分布,后随机分布或均匀分布。河滩生境中刺萼龙葵聚集分布不明显,可能是由于河滩生境与其他生境相比,水分更充足,土壤相对肥沃,刺萼龙葵种群内对自身生长所需资源的竞争相对较弱,种群主要呈现随机分布。此外,在河滩生境中,仅中型植株表现出微弱的聚集性,而在草原生境中,大型植株分布表现出较强的聚集性,在荒地生境中,3种植株类型均表现出一定的聚集分布。在草原和荒地生境中,大型植株和中型植株在中小尺度范围内表现出聚集性,且大型植株相对于中型植株表现出更强的聚集性。由于草原生境和荒地生境的土壤相对干旱,且荒地土壤相对贫瘠,不利于刺萼龙葵种群繁殖和扩张,在资源有限的情况下,聚集分布可能更有利于刺萼龙葵种群占据更多生态位,以获得更多资源,增强同其他物种的竞争力。由此可推断,在有利的环境下,刺萼龙葵分布随机性较大,而在不利的环境下,刺萼龙葵有由中心向四周爆发的趋势,且越靠近中心位置,越呈现出明显的聚集分布。
梅军林等(2017)关于红背山麻杆种群的点格局研究表明,空间正关联或负关联主要发生在小尺度范围内,且空间关联性随研究尺度的增大而减弱,最终趋于无相关性。本研究中,在荒地生境的中小尺度范围内,刺萼龙葵的大、中、小3种植株类型间均表现出一定的正相关性,且随着尺度的增大,不断趋于无相关性或负相关;草原生境中,在小尺度范围内,大、中、小3种植株类型间均表现出一定的正相关性,且随着尺度的增大,不断趋于无相关性。由此可推测以上2种生境中的刺萼龙葵种群演替规律,即在小尺度内,刺萼龙葵大、中、小个体间存在一定的相互有利生态关系,以利于其种群的不断扩大,随着种群的不断扩大,在大尺度内,种群内个体间互不影响,或表现出一定的竞争关系。河滩生境中,仅中型植株与小型植株间在小尺度表现出正相关性,大、中、小3种植株类型间随着尺度的增大,不断趋于无相关性或负相关。河滩生境相较于草原生境和荒地生境,土壤水分和养分等资源充足,刺萼龙葵个体间独立性较强,关联较小;与大型植株相比,小型植株和中型植株对生长所必需资源的竞争力微弱,幼小型植株主要出现在较大型植株的斑块间隙,其分布在很大程度上取决于较大型植株为其创造的有利环境(樊登星和余新晓,2016),在长期自然选择过程中形成的生活对策是小型植株与较大型植株间通过相互庇护,发挥群体效应,以增大个体存活的机会(张金屯和孟东平,2004)。由此推断,由于刺萼龙葵种群内个体间缺乏激烈的竞争,因此更加剧其种群的大面积暴发。
4 结论
入侵植物刺萼龙葵的群落空間格局在不同资源环境条件下均呈现有利于种群扩散的分布特征,且不同株型间易形成互利的生态关系,是其成功入侵的主要驱动因素之一。
参考文献:
樊登星,余新晓. 2016. 北京山区栓皮栎林优势种群点格局分析[J]. 生态学报,36(2):318-325. [Fan D X,Yu X X. 2016. Spatial point pattern analysis of Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing[J]. Acta Ecologica Sinica,36(2):318-325.]
关广清. 2000. 杂草种子图鉴[M]. 北京:科学出版社. [Guan G Q. 2000. Graphics of weed seeds[M]. Beijing: Science Press.]
胡砚秋,李文斌,崔佳玉,苏志尧. 2016. 亚热带常绿阔叶林优势种个体及生物量的点格局分析[J]. 生态学报,36(4):1066-1072. [Hu Y Q,Li W B,Cui J Y,Su Z Y. 2016. Spatial point patterns of dominant species by individualtrees and biomass in a subtropical evergreen broad-leaved forest[J]. Acta Ecologica Sinica,36(4):1066-1072.]
李国春,宋华东,李琦,卜书海. 2017. 太白山巴山冷杉林主要树种与开花秦岭箭竹的空间点格局分析[J]. 应用生态学报,28(11):3487-3493. [Li G C,Song H D,Li Q,Bu S H. 2017. Spatial point pattern analysis of main trees and flowering Fargesia qinlingensis in Abies fargesii forests in Mt. Taibai of the Qinling Mountains,China[J]. Chinese Journal of Applied Ecology,28(11):3487-3493.]
李明辉,何风华,刘云,潘存德. 2003. 林分空间格局的研究方法[J]. 生态科学,22(1):77-81. [Li M H,He F H,Liu Y,Pan C D. 2003. Analysis methods of stand spatial distribution pattern[J]. Ecological Science,22(1):77-81.]
梅军林,庄枫红,马姜明,覃扬浍,梁士楚,姜勇. 2017. 桂林喀斯特地区克隆生长红背山麻杆种群的点格局分析[J]. 生态学报,37(9):3164-3171. [Mei J L,Zhuang F H,Ma J M,Qin Y H,Liang S C,Jiang Y. 2017. Spatial point pattern analysis of Alchornea trewioides population clonal growth in the karst area of Guilin[J]. Acta Ecologica Sinica,37(9):3164-3171.]
时培建,郭世权,杨清培,王兵,杨光耀,方楷. 2010. 毛竹的异质性空间点格局分析[J]. 生态学报,30(16):4401-4407. [Shi P J,Guo S Q,Yang Q P,Wang B,Yang G Y,Fang K. 2010. Inhomogeneous spatial point pattern analysis of moso bamboo(Phyllostachys edulis)[J]. Acta Ecologica Sinica,30(16):4401-4407.]
王梅,赵晨光,王莹,楚光明. 2015. 盐穗木种群空间点格局对地下水埋深的响应[J]. 林业科学,51(11):17-24. [Wang M,Zhao C G,Wang Y,Chu G M. 2015. Response of spatial point pattern of Halostachys caspica population tounderground water depth[J]. Scientia Silvae Sinicae,51(11):17-24.]
王瑞,唐瑶,张震,万方浩. 2018. 外来入侵植物刺萼龙葵在我国的分布格局与早期监测预警[J]. 生物安全学报,27(4):284-289. [Wang R,Tang Y,Zhang Z,Wan F H. 2018. The distribution pattern and early monitoring for preventing further expansion of Solanum rostratum in China[J]. Journal of Biosafety,27(4):284-289.]
杨洪晓,张金屯,吴波,李晓松,张友炎. 2006. 毛乌素沙地油蒿种群点格局分析[J]. 植物生态学报,30(4):563-570. [Yang H X,Zhang J T,Wu B,Li X S,Zhang Y Y. 2006. Point pattern analysis of Artemisia ordosica population in the Mu Us sandy land[J]. Journal of Plant Ecology,30(4):563-570.]
杨云方,丁晖,徐海根,方炎明,陈晓,伊贤贵,徐辉,杨青,温小荣,徐鲜均. 2013. 武夷山典型常绿阔叶林4个主要植物种群点格局分析[J]. 生态与农村环境学报,29(2):184-190. [Yang Y F,Ding H,Xu H G,Fang Y M,Chen X,Yi X G,Xu H,Yang Q,Wen X R,Xu X J. 2013. Point pattern analysis of four main plant populations in evergreen broad-leaved forests typical of Wuyi mountains,Southeast China[J]. Journal of Ecology and Rural Environment,29(2):184-190.]
張金屯. 1998. 植物种群空间分布的点格局分析[J]. 植物生态学报,22(4):344-349. [Zhang J T. 1998. Analysis of spatial point pattern for plant species[J]. Acta Phytoecologica Sinica,22(4):344-349.]
张金屯,孟东平. 2004. 芦芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报,24(1):35-40. [Zhang J T,Meng D P. 2004. Spatial pattern analysis of individuals in different age-classes of Larix principis-rupprechtii in Luya mountain reserve,Shanxi,China[J]. Acta Ecologica Sinica,24(1):35-40.]
张丽娟,娄安如. 2018. 入侵植物刺萼龙葵的繁殖保障及其与种群大小的关系[J]. 北京师范大学学报(自然科学版),54(4):491-497. [Zhang L J,Lou A R. 2018. Reproductive assurance in invasive Solanum rostratum and its relationship to population size[J]. Journal of Beijing Normal University(Natural Science),54(4):491-497.]
赵欣胜,崔保山,孙涛,吕卷章,路峰. 2011. 不同生境条件下中国怪柳空间分布点格局分析[J]. 生态科学,30(2):142-149. [Zhao X S,Cui B S,Sun T,Lü J Z,Lu F. 2011. Analysis of spatial point pattern of Tamarix chinensis in different habitats[J]. Ecological Science,30(2):142-149.]
钟艮平,沈文君,万方浩,王进军. 2009. 用GARP生态位模型预测刺萼龙葵在中国的潜在分布区[J]. 生态学杂志,28(1):162-166. [Zhong G P,Shen W J,Wan F H,Wang J J. 2009. Potential distribution areas of Solanum rostratum in China:A prediction with GARP niche model[J]. Chinese Journal of Ecology,28(1):162-166.]
Diggle P J. 2003. Statistical analysis of spatial point patterns[M]. The 2nd Edition. London:Hodder Education Publi-shers.
Druckenbrod D L,Shugart H H,Davies I. 2005. Spatial pa-ttern and process in forest stands within the Virginia piedmont[J]. Journal of Vegetation Science,16(1):37-48.
Horvitz N,Wang R,Wan F H,Nathan R. 2017. Pervasive human-mediated large-scale invasion:Analysis of spread patterns and their underlying mechanisms in 17 of Chinas worst invasive plants[J]. Journal of Ecology,105(1):85-94.
Li W,Zhang G F. 2015. Population structure and spatial pa-ttern of the endemic and endangered subtropical tree Parrotia subaequalis(Hamamelidaceae)[J]. Flora-Morphology,Distribution,Functional Ecology of Plants,212:10-18.
Nguyen H H,Uria-Diez J,Wiegand K. 2016. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam[J]. Journal of Vegetation Science,27(2):318-327.
Ripley B D. 1976. The second-order analysis of stationary point processes[J]. Journal of Applied Probability,13(2):255-266.
Ripley B D. 1977. Modeling spatial patterns[J]. Journal of the Royal Statistical Society(Series B),39(2):172-212.
Wiegand T,Moloney K A. 2013. Handbook of spatial point-pattern analysis in ecology[M]. Boca Raton:CRC Press.
Wilschut L I,Laudisoit A,Hughes N K,Addink E A,de Jong S M,Heesterbeek H A P,Reijniers J,Eagle S, Dubyanskiy V M,Begon M. 2015. Spatial distribution patterns of plague hosts:Point pattern analysis of the burrows of great gerbils in Kazakhstan[J]. Journal of Biogeography,42(7):1281-1292.
猜你喜欢
湖北农业科学(2018年3期)2018-03-27
现代农业科技(2018年1期)2018-02-03
绿色科技(2017年16期)2017-09-22
吉林农业(2017年8期)2017-08-08
青年时代(2017年16期)2017-07-03
科学与财富(2017年17期)2017-06-16
南方农业·下旬(2017年1期)2017-05-31
江苏农业科学(2016年1期)2017-05-17
中国科技纵横(2016年15期)2016-12-29
绿色科技(2014年9期)2014-10-30
