
P38 MAPK信号通路在金黄色葡萄球菌诱导U937细胞凋亡中的作用
2020-04-13 14:18肖楚丽
国际感染病学(电子版) 2020年3期
肖楚丽
邵阳学院,湖南 邵阳 422000
金黄色葡萄球菌(SA)属于化脓性球菌的一种类型,也是造成院内感染的主要原因,一旦患者出现感染,可能引发食物中毒、假膜性肠炎以及烫伤样皮肤综合征,对住院患者的治疗造成非常大的威胁。金黄色葡萄球菌能调节人体的逃避宿主免疫防御系统,直至死亡,但是,其引发死亡的作用机制并不明确,所以,研究金黄色葡萄球菌中细胞走向凋亡的重要介导因子,具有重要的意义,也是目前临床研究的一个重要方向。目前,仅有少数学者发现,可能与丝裂原活化蛋白激酶(MAPK)活性高低有关,在信号通路的传导中,有着重要的促进作用,但是,MAPK是如何通过信号通路,产生作用,其作用机制和原理,还有待深入的分析,所以,对P38MAPK信号传导通路诱导金黄色葡萄糖球菌,至细胞凋亡的作用进行分析,是一项重要的研究课题[1]。本文对P38 MAPK信号通路在金黄色葡萄球菌诱导U937细胞凋亡中的作用进行分析,内容如下:
1 材料与方法
1.1 材料 100株金黄色葡萄球菌均自实验室中获取,菌株由实验室提供,并在不含有抗生素的5%小牛血清RPMI1640培养基重悬并进行稀释。试剂选择:P38 MAPK、PHOSPHO-P38MAPK抗体,P38 MAPK抑制剂,凋亡试剂盒、细胞培养试剂。
1.2 方法
①细胞培养以及抑制剂加入。将U937(人巨噬细胞)中置入10%小牛血清,与2mmol/L L-谷氨酸钠、100U/ml青霉素、100g/ml链霉素放于RPMI1640培养基中,进行培养箱培养,温度为37℃,并在SA感染前1h加入不同浓度的P38 MAPK抑制剂。
②SA感染U937细胞。感染前需要使用不含抗生素的RPMI1640培养基将U937进行细胞的清洗并进行重悬,调整浓度,细胞与细菌的比例为1:20,之后分别培养0/15/30/60min,连续2次进行离心处理,将未吞没的细菌标本清洗干净,将其放入青霉素、链霉素清RPMI1640的培养皿中进行培养,放置3h后,进行感染细胞提取,对其进行定性分析。
连续离心2次,清洗未吞噬的细菌标本,放置于2倍小牛血清、青霉素以及链霉素的清RPMI1640培养基中,3h后提取感染的U937细胞,观察变化。
③双染分析。使用流式细胞分析仪,将106细胞加入含有an-nexinV的缓和液中,缓和液剂量为60µL,在4℃温度下避光15分钟,注意区分早期凋亡细胞、晚期凋亡细胞以及坏死细胞,并在实施检查之前同时染an-nexinV与PI,结合测试仪分析结果。
④Western免疫印迹。在U937细胞处理过一段时间后,实施PBS洗涤,在3次后将其中裂解液进行重悬,时间为20min,之后使用同样数量的蛋白12%的SDS-聚丙烯酰胺凝胶电泳,使用蛋白转印系统将蛋白转印至膜上,60min后使用5%奶粉在三羟甲基氨基甲烷缓冲盐水中进行封闭,并按照说明书的操作要求,对其进行Tween20洗涤,经过底物显色或放射自显影以检测电泳分离的特异性目的基因表达的蛋白成分。
1.3 观察指标 对比U937细胞凋亡百分比在不同时间段的表现。
1.4 统计学处理 本次研究数据均采用统计学软件SPSS20.0进行处理,计数资料采用χ2表示,P检验,P<0.05具有统计学意义。
2 结果
随着时间的增加,U937细胞凋亡百分比显著提升,详见表1。
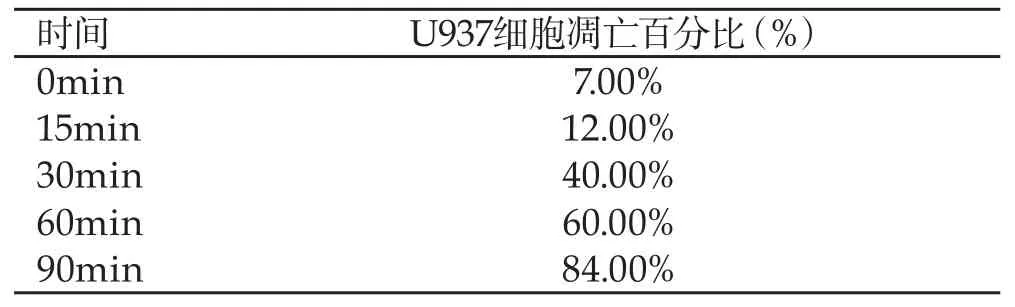
表1 对比不同时间下U937细胞凋亡百分比
3 讨论
金黄色葡萄糖球菌的抵抗力是最强的,属于一种无芽胞菌,在人体中大量的存在,其生存条件简单,如皮肤表面附着,鼻腔内部附着等。研究显示,MAPK的存在途径非常多样化,被学者知晓的目前就有4种,P38 MAPK信号通路是其中一种,主要通过丝氨酸以及苏氨酸同时磷酸化之后出现激活,因此需要对该方面进行分析。根据学者研究指出,p38MAPK可介导、分化细胞的分化和增殖,并且大量的存在于对中生物学效应中,有时候,甚至可以产生抑制等相反的作用,但是,其具有很强的细胞依赖性,在致病性细菌侵袭的细胞中,p38MAPK信号通路均表现出很强的活跃性。例如,p38MAPK在巨噬细胞中,能参与脂多糖(LPS)的活化活动,或者是大肠杆菌感染上皮细胞中,p38MAPK在李斯特菌、沙门菌等致病菌中,能借助这些病菌的诱导作用,p38MAPK被活化。而根据学者研究发现,在大肠杆菌标准株ATC25922能诱导U937细胞逐渐走向凋亡,而此时p38MAPK磷酸化水平也随着时间的延长,而逐渐增长,该学者研究表示,p38MAPK在U937细胞凋亡过程中,发挥着重要的作用。所以,对于金黄色葡萄球菌中p38MAPK是否也发挥着同样的作用,是一个值得深入研究的问题。
而通过本文研究,也再次证实随着时间的增加,U937细胞凋亡百分比显著提升。分析原因:丝裂原激活的蛋白激酶(MAPKS)又称为丝氨酸-苏氨酸蛋白激酶,介导信号自细胞表面向核内传递,属于一种信号系统,在细胞的生长、增殖、分化以及凋亡的过程中起到非常重要的作用,而P38 MAPK属于MAPKS的组成部分,主要介导炎症、应激、损伤等信号的传递,可以被炎细胞因子、毒素等应激因素激活,因此其在调节炎症反应等方面发挥的作用较为显著,同时P38通路异常过度激活可能导致细胞正常停滞或者凋亡;而激活P38 MAPK的原因可能是因为患者受到应激性的刺激,或病理性损伤,引发介导细胞死亡,因此一些细胞可在P38 MAPK的介导下出现分化以及增殖,并且在活化之后生物效应多样化显著,有些甚至与原本属性相反,可能存在依赖性,此时,研究P38MAPK信号通路在金黄色葡萄糖球菌中的诱导作用,有着重要的意义。
通过研究发现,金黄色葡萄球菌诱导U937细胞死亡时,其细胞的死亡率,是随着时间延长而逐渐升高的,可见金黄色葡萄球菌可以将P38 MAPK激活,因此在进行该疾病预防的过程中需要注意对该方面的预防,为细菌的控制提供更加理想的条件[7];对P38 MAPK进行研究发现,其可以被吞噬细胞中的脂多糖成分逐渐活化,最终释放大量的沙门菌或里斯特菌,附着在大肠上,引起病变,最终通过P38MAPK产生诱导作用,整个过程中完全遏制了TNF-a的活性,最终使得人中性粒细胞IL-8发生超氧化离子,发生MIP诱导化学趋化性,介导P38 MAPK活化,进而启动信号传导系统,促进细胞表达、合成以及释放IL-1、ILK-6等多种炎性因子,参与至炎性反应的过程中,而U937细胞在感染金黄色葡萄球菌后P38 MAPK的活性,发现其在15min后开始升高,证明其具有激活的效果,因此可以降低抑制剂依赖性细胞的凋亡,为疾病的控制提供条件[8-9]。
综上分析,P38 MAPK信号通路是金黄色葡萄球菌诱导U937细胞凋亡的重要传导。
猜你喜欢
食品安全导刊(2021年20期)2021-08-30
中华养生保健(2020年3期)2020-11-16
实验与检验医学(2019年2期)2019-04-15
兽医导刊(2019年1期)2019-02-21
创新作文(1-2年级)(2018年2期)2018-09-13
创新作文(小学版)(2018年4期)2018-07-06
学习报·教育研究(2017年33期)2017-08-31
现代检验医学杂志(2016年2期)2016-11-14
现代检验医学杂志(2016年2期)2016-11-14
中外医疗(2015年18期)2016-01-04
