
海南三亚和黄淮海地区玉米小斑病菌致病性及遗传多样性分析
2020-04-13 05:49常佳迎刘树森石洁郭宁张海剑马红霞杨春凤
中国农业科学 2020年6期
常佳迎,刘树森,石洁,郭宁,张海剑,马红霞,杨春凤
海南三亚和黄淮海地区玉米小斑病菌致病性及遗传多样性分析
常佳迎,刘树森,石洁,郭宁,张海剑,马红霞,杨春凤
(河北省农林科学院植物保护研究所/农业农村部华北北部作物有害生物综合治理重点实验室/河北省农业有害生物综合防治工程技术研究中心,河北保定 071000)
【目的】明确海南三亚玉米南繁基地和黄淮海玉米主产区的玉米小斑病菌()菌株的致病性差异,以及遗传多样性和亲缘关系的远近,为海南三亚南繁基地育种过程中玉米品种的抗病性筛选提供理论依据,并对玉米品种在黄淮海地区的推广种植提供参考。【方法】对采集自海南三亚和黄淮海地区的玉米小斑病病叶进行病原菌分离,经过形态学和分子生物学方法对分离到的菌株进行鉴定,共获得61株玉米小斑病菌菌株。采用人工喷雾接种法对61株小斑病菌菌株的致病力进行鉴定;并选取16条扩增条带清晰、重复性好、多态性高的引物进行ISSR-PCR扩增,利用Popgen32生物软件计算群体间的遗传距离和遗传相似性,并利用NTsys2.10e软件进行聚类分析,构建海南三亚与黄淮海地区小斑病菌的聚类图。【结果】致病性测定结果表明,海南三亚地区未分离到弱致病力菌株,强致病力菌株个数占三亚地区总菌株的57.1%;黄淮海地区分离到的弱致病力菌株占黄淮海地区总菌株的47.5%,强致病力菌株占黄淮海地区总菌株的17.5%。遗传多样性分析结果表明,在群体平均水平上,Nei’s基因多样性指数(H)为0.2820,Shannon’s信息指数(I)为0.4197,表明玉米小斑病菌具有一定的遗传变异;不同地理种群间的遗传相似系数为0.9028—0.9618,遗传距离为0.0390—0.1023,表明整体遗传相似系数较高,遗传距离较近;但不同地理种群间仍存在不同程度的遗传分化,其中,河南与河北的菌株群体遗传相似系数最高(GS=0.9618),亲缘关系最近;河南和三亚地区的菌株群体遗传相似系数最低(GS=0.9028),亲缘关系相对较远。聚类分析结果表明,三亚地区菌株和黄淮海地区菌株在相似系数为0.722时,明显被分为两大类群。【结论】黄淮海玉米主产区与海南三亚玉米南繁基地的玉米小斑病菌在致病力上存在一定差异,三亚地区强致病力菌株的分离频率高于黄淮海地区。黄淮海地区与三亚地区的菌株群体具有丰富的遗传多样性,且遗传多样性与地理来源有一定关系,地域相邻的菌株遗传关系较近;但三亚地区与黄淮海地区的菌株遗传相似系数相对较高,亲缘关系较近,且两种群间存在一定基因交流。因此,三亚地区作为玉米的南繁基地,其自然发病条件下筛选出对小斑病具有抗性的玉米品种或抗性亲本材料,对黄淮海地区小斑病抗性的逐年提高具有一定的影响,有利于减轻黄淮海地区病害的发生。
玉米小斑病菌;玉米小斑病;南繁基地;海南三亚;黄淮海地区;致病性;遗传多样性
0 引言
【研究意义】玉米是中国主要的粮食作物之一,具有很高的营养价值,也是重要的饲料作物和工业原料,其经济地位不容忽视。随着玉米种植面积的逐年增长,栽培模式向轻简化发展,生产上对抗病品种的需求日益迫切。目前,有能力采用人工接种技术选育抗病品种的单位和育种家较少,在常年发病较重区域设置自然鉴定圃是育种家的首选。海南省三亚市作为玉米育种扩繁加代的一个重要地区,其小斑病发生较为普遍。能否依据品种(系)在海南田间小斑病的自然发病结果,淘汰抗性差的品种(系),选择抗性较好的品种(系),并在黄淮海夏玉米种植区应用是一个亟待解决的问题。因此,明确三亚地区与黄淮海地区病原菌致病性和遗传多样性上的差异,对育种过程中玉米品种小斑病抗性筛选及其在黄淮海地区种植具有重要意义。【前人研究进展】玉米小斑病是黄淮海和南方玉米种植区主要的叶部病害之一,其致病菌为玉蜀黍平脐孺孢(),可在玉米全生育期侵染引起病症,具有较强的侵染性和致病力[1-3]。在适宜的条件下,玉米小斑病菌可在接种24 h后侵染引起叶片坏死[4],随后在病斑上产生的分生孢子可借助气流传播,成为再侵染源[5]。玉米小斑病的发生和流行严重影响了玉米的产量和品质,发病较轻的地块可造成10%—30%的产量损失,重病田减产高达80%[6-8]。研究表明,小斑病菌不同菌株在菌丝生长速率、产孢量、菌落特征等生物学特性方面存在差异[9-10];不同菌株对相同玉米品种的致病性存在明显差异[1-3]。简单序列重复区间扩增多态性(inter-simple sequence repeat,ISSR)分子标记技术具有操作简单、稳定性好、多态性高等优点[11-12],目前广泛应用于病原菌的遗传多样性分析,如玉米灰斑病菌(spp.)[13]、玉米大斑病菌()[14]、玉米圆斑病菌()[15]等。郭云燕等[16]利用ISSR分子标记技术对不同省份间的玉米南方锈病病原菌()进行分析,探讨了其病原菌初侵染来源;马红霞等[17]采用ISSR-PCR方法对不同地区的禾谷镰孢()复合种进行了遗传多样性分析,结果表明不同地理种群间遗传多样性与地理来源有关;常佳迎等[18]采用ISSR分子技术分析了黄淮海地区不同年份的弯孢叶斑病菌(),结果表明年度间菌株具有一定差异,推测菌株的初侵染源为外来菌源;梅玉云[19]和廖蕾[20]采用ISSR分子标记技术分别对安徽省不同地区和福建省不同地区的小斑病菌进行遗传多样性分析,结果表明小斑病菌群体存在丰富的遗传多样性,同一省份内不同地区菌株的亲缘关系与地理来源有一定的相关性。【本研究切入点】玉米小斑病菌具有较强的寄主适应性和致病力,且来自同一省份不同地区的菌株致病性和遗传多样性存在差异,但海南三亚地区与黄淮海地区的病原菌致病性和亲缘关系并未明确报道。【拟解决的关键问题】对采集自海南三亚和黄淮海地区的玉米小斑病菌菌株进行致病性和遗传多样性差异分析,了解两地菌株的致病性差异和亲缘关系的远近,明确三亚地区自然发病条件下筛选获得的玉米材料在黄淮海地区是否具有种植价值,以期为周年内两代抗病育种工作的顺利进行提供参考。
1 材料与方法
试验于2018年10月至2019年6月在河北省农林科学院植物保护研究所完成。
1.1 材料
供试菌株:2018年9月从河北、河南和山东省采集夏玉米小斑病病叶,2019年2月从海南省三亚市采集南繁玉米小斑病病叶,放置于标本夹中,待水分去除后转入干燥信封袋中保存备用。采用常规病害组织分离法[21]和单孢纯化法[22]获得病原菌,对获得的病原菌进行形态学和分子生物学鉴定(ITS序列分析),共获得61株玉蜀黍平脐蠕孢(表1)。
供试品种:选用小斑病感病自交系B73进行病原菌致病性测定。
1.2 致病性分析
1.2.1 病原菌扩繁 将分离获得的61个菌株分别转接至PDA培养基平板上,黑暗条件下30℃培养5 d后,转接到高粱粒培养基上培养10 d,用无菌水将分生孢子洗下,制成分生孢子悬浮液(1×105个/mL),加入0.01%吐温20(V/V)备用。
1.2.2 室内致病性测定 在花盆(上口直径:11.5 cm,下口直径:10.5 cm,高:8.5 cm)中种植玉米自交系B73,每盆种植4株,共种植61盆,待玉米苗长至3叶期,采用人工喷雾接种的方法,喷施分生孢子悬浮液,每个菌株喷施1盆,且均匀喷施等量(10 mL)的分生孢子悬浮液。接菌后保湿48 h,7 d后调查发病情况,测量病斑大小,记录病斑特征,采用透明刻度板(5 mm方格)评估发病面积。
1.3 遗传多样性分析
1.3.1 模板DNA的制备 将分离获得的61个菌株分别接种到PDA培养基上,30℃培养7 d,收集气生菌丝。采用真菌基因组提取试剂盒(北京艾德来生物科技有限公司),按照说明书提取各菌株的DNA。采用ND2000微量紫外分光光度计(Thermo,USA)测定所获得的DNA质量和浓度,统一稀释至30 ng·μL-1,于-20℃保存备用。
1.3.2 ISSR分析 根据梅玉云[19]和廖蕾[20]的研究,选取16条ISSR引物用于本试验(表2)。ISSR反应体系:DNA模板1 μL(30 ng·μL-1),2×EsTaq Master Mix 10 μL,Primer(10 μmol·L-1)1.5 μL,加ddH2O补足至20 μL。PCR反应程序:95℃预变性5 min,94℃变性40 s,退火(视不同引物而定)45 s,72℃延伸1.5 min,30个循环,最后72℃延伸10 min。
PCR扩增产物在1.8%琼脂糖凝胶上以100 V,电流100 mA电泳55 min,使用GelDoc XR+凝胶成像系统照相。
1.3.3 数据分析 统计并记录ISSR电泳图谱条带,在相同大小的位置有条带的标记为1,无条带的标记为0。采用Popgen32软件进行数据分析,计算海南三亚和黄淮海地区群体间的Shannon’s多样性指数、群体间的遗传距离和遗传相似性[23-24];利用NTsys2.10e软件,采用clustering模块中的SAHN按照非加权组平均法(UPGMA)进行聚类分析,并构建系统聚类图。
2 结果
2.1 不同地区小斑病菌的致病性分析
致病性测定结果表明,61株小斑病菌在玉米自交系B73上的致病力、病斑大小和病斑形状存在明显差异。病斑形状可分为:受叶脉限制的长方形和不受叶脉限制的椭圆形,有明显褐色边缘和无褐色边缘;病斑颜色分为浅黄色、黄褐色和深褐色(表1)。依据《中华人民共和国农业行业标准——玉米抗病虫性鉴定技术规范(NY/T1248.2—2016)》的分级标准,并根据发病面积将致病力划分为强、中、弱3个等级,发病面积≤10%为弱致病力菌株;10%<发病面积≤30%为中等致病力菌株;发病面积>30%为强致病力菌株。由表1可见,黄淮海地区40个菌株中有19个为弱致病力菌株,占黄淮海地区总菌株的47.5%,14个菌株为中等致病力菌株,占黄淮海地区总菌株的35.0%,7个为强致病力菌株,占黄淮海地区总菌株的17.5%;海南三亚地区的21个菌株中无弱致病力菌株,9个中等致病力菌株,占三亚地区总菌株的42.9%,其余12个菌株为强致病力菌株,占三亚地区总菌株的57.1%。观察结果表明,在玉米品种上形成的病斑颜色深、无褐色边缘的菌株致病力普遍较强。

表1 黄淮海地区和海南三亚小斑病菌的致病性测定

续表1 Continued table 1
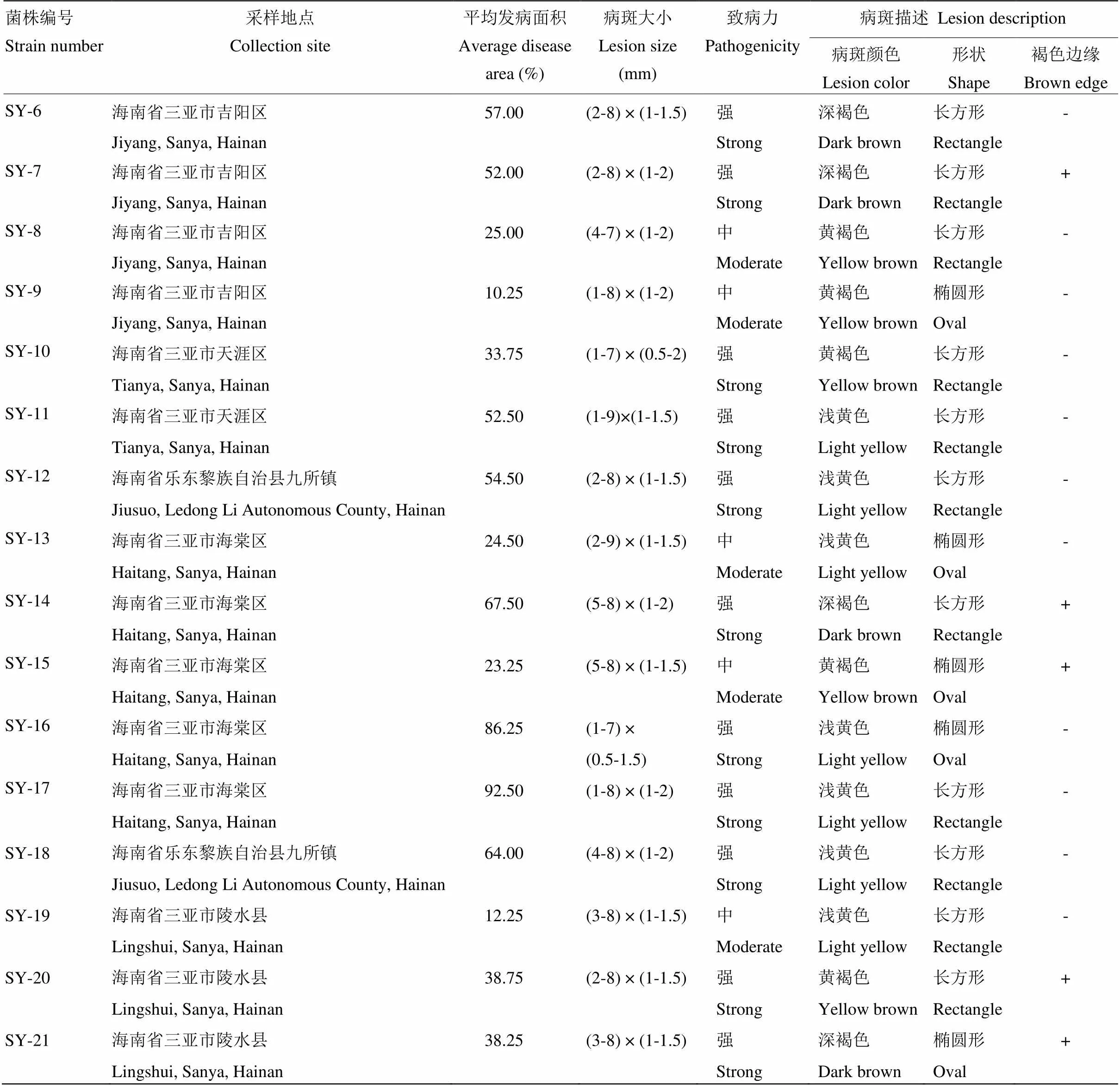
续表1 Continued table 1
“-”:无褐色边缘no brown edge;“+”:有褐色边缘With brown edges
2.2 不同地区小斑病菌的遗传多样性
2.2.1 不同地区小斑病菌的遗传多态性分析 选取的16条引物具有较好的多态性,共获得177个条带,分布在100—10 000 bp,其中多态性条带140个,占79.10%。不同引物扩增获得的条带数目为7—17条,平均每个引物获得9个多态性条带,其中引物808获得条带数最多(17条),引物812获得的条带数最少,仅为7条;16条引物的多态性条带比率为44.44%—100.00%(表2)。
将61株小斑病菌按省份分为4个地理种群,其遗传多态性分析如表3所示。在群体平均水平上,多态性位点为140个,多态性位点平均比率为79.10%,观察等位基因数(Na)为1.7910,有效等位基因数(Ne)为1.4904,Nei’s基因多样性指数(H)为0.2820,Shannon’s信息指数(I)为0.4197,表明玉米小斑病菌具有一定的遗传变异。
不同地理来源的种群间,H值在0.1717—0.2783,I值在0.2505—0.4072,表明不同地理种群间的玉米小斑病菌遗传多样性具有一定差异。总体而言,河北(0.2783)和海南三亚(0.2437)种群遗传多样性较高,河南(0.2137)种群次之,山东(0.1717)种群较低(表3),基因多样性指数的差异可能受分析所用种群中菌株数量的影响。
分析得到4个地理种群总基因多样性(Ht)为0.2664,各地理种群内基因多样性(Hs)为0.2268,地理群体间的基因多样性(Dst)为0.0396,表明相同省份来源的菌株具有较近的亲缘关系。地理种群间的基因分化系数(Gst)为0.1484,这说明不同地理种群间遗传变异占种群总遗传变异的14.84%,大部分遗传变异(85.16%)发生在种群内。根据遗传分化系数估算的基因流的值Nm=2.8690(>1),表明地理种群间存在一定的基因交流。
2.2.2 不同地区小斑病菌的遗传相似性分析 为进一步分析玉米小斑病菌各地理种群间的遗传分化程度,将分离到的61株小斑病菌按省份分组,计算Nei’s遗传相似性(GS)和遗传距离(GD)(表4)。不同地理种群间的遗传相似系数为0.9028—0.9618,遗传距离为0.0390—0.1023。遗传相似系数均>0.9000,遗传距离最大仅为0.1023,表明整体遗传相似系数较高,遗传距离较近;但不同地理种群间仍存在不同程度的遗传分化。其中,河南与海南的菌株群体遗传相似系数相对较低(GS=0.9028),遗传距离较远(GD=0.1023),亲缘关系相对较远;河北与河南的遗传相似系数相对较高(GS=0.9618),遗传距离相对较近(GD=0.0390),亲缘关系较近。

表2 ISSR引物扩增结果
2.2.3 不同地区菌株间亲缘关系的聚类分析 利用NTsys2.10e软件对61株小斑病菌进行ISSR聚类分析,菌株间具有丰富的遗传多样性,相似系数在0.71—0.95(图1)。在相似系数为0.722时,所有的菌株被分成两大群(Group 1和Group 2)。在相似系数为0.74时,Group 1可分为2个亚群,subgroupⅠ包含1个河南菌株,2个三亚菌株,13个河北菌株;subgroupⅡ包含2个山东菌株,2个三亚菌株,9个河南菌株,11个河北菌株。Group 2包含1个山东菌株,3个河北菌株,17个三亚菌株。根据ISSR聚类分析结果,黄淮海地区的菌株多分布于Group 1,而三亚地区的菌株群体多分布于Group 2。

图1 玉米小斑病菌的UPGMA聚类分析

表3 玉米小斑病菌不同地理种群的遗传多样性水平

表4 玉米小斑病菌不同地理种群间的遗传相似性和遗传距离
对角线上方为遗传相似系数,下方为遗传距离Nei’s genetic similarity coefficient (above diagonal) and genetic distance (below diagonal)
3 讨论
研究表明,玉米小斑病菌致病力分化十分明显[25],且小斑病菌的毒力与地理位置无关[26],不同菌株对相同玉米品种/自交系的致病力不同,且形成的病斑类型均有明显差异[1-3]。在中国玉米产区具有强致病力和中等致病力的小斑病菌菌株占有明显优势[27-28]。本研究结果表明,不同菌株在同一个玉米自交系上造成的病斑类型不同,病斑大小也存在一定差异。海南三亚地区未分离到弱致病力菌株,强致病力菌株占三亚地区总菌株的57.1%,为优势种群;黄淮海地区分离到的弱致病力菌株占黄淮海地区总菌株的47.5%,为主要种群,强致病力菌株占17.5%,因此海南三亚地区强致病力菌株的分离频率高于黄淮海地区。推测可能是受黄淮海玉米种植区推广抗病品种的影响,导致病原菌的致病力减弱;而在海南三亚地区进行材料的南繁,不考虑材料的抗病性,其南繁材料中大部分对小斑病表现更为敏感,因此在三亚也聚集了更多的强致病力菌株。
本研究通过对分离自黄淮海地区和海南三亚地区的61株玉米小斑病菌的ISSR片段多样性进行分析,共获得140个多态性条带,多态性比率为79.10%,表明黄淮海地区和三亚地区的小斑病菌具有丰富的遗传变异,不同地理种群间小斑病菌的遗传多样性具有一定差异,这与梅玉云[19]和廖蕾[20]的研究结果一致。不同地理种群间的遗传相似系数和遗传距离表明,整体遗传相似系数较高,遗传距离较近;但地理种群间存在地域相邻,遗传关系相对较近,遗传相似系数较高的特点。聚类分析显示,海南三亚与黄淮海地区的小斑病菌具有一定地理隔离,且小斑病菌的遗传亲缘关系与地理来源有一定的相关性;但Group 1中包含4个三亚地区的菌株而Group 2包含4个黄淮海地区的菌株,同时遗传分化系数估算的基因流的值Nm=2.8690(>1),表明黄淮海和海南三亚地区地理种群间存在一定的基因交流,可能是在育种材料的调运过程中随种子、穗轴苞叶等传播,以及北上气流的影响,导致部分菌株在三亚和黄淮海地区的交叉定殖。鉴于海南的小斑病菌致病力更强,今后应加强该病害的检疫和监测工作。
王群[29]对299份山东省玉米引种备案品种(系)和101份黄淮海玉米趋势试验品种(系)进行玉米小斑病的抗性鉴定,结果显示表现为高抗和抗病的品种占96.25%,没有表现高感的玉米品种;施艳等[30]鉴定了2015—2017年河南省夏玉米品种对小斑病的抗性,结果表明中抗品种占主要地位,而具有抗性的品种比率呈逐年上升趋势,且同一品种在不同年份抗性表现较为稳定;陆银萍[31]采用人工喷雾法检测了安徽省2010—2014年参与鉴定的696个玉米品种对小斑病的抗性,发现抗性等级为高抗和中抗的品种占鉴定总数的比例呈总体上升趋势;由此可见,黄淮海地区近年来的主栽品种对小斑病的抗性提高,可能得益于育种过程中对品种抗病性的筛选。同时,建议育种家对已经形成的有望生产应用的组合在海南三亚地区开展小斑病抗性鉴定工作,也可在品种选配中采用在三亚表现抗小斑病的亲本,在较高压力下确定新品种的抗性水平,组配出抗小斑病的品种。
海南三亚作为玉米南繁育种的重要基地,其小斑病发生普遍,本研究结果证明三亚地区强致病力菌株所占比例较黄淮海地区高,三亚地区菌株与黄淮海地区菌株的整体遗传相似系数较高,遗传距离较近,且两地之间的菌株存在一定的基因交流。因此,在三亚地区田间育种过程中,根据田间自然发病情况对品种的抗病性进行筛选具有一定意义,有利于筛选出对小斑病抗性较好的玉米品种。因黄淮海地区弱致病菌群占主要地位,因此对于在海南田间小斑病发生较重的玉米品种(系),若其他田间性状(如产量等)表现较好,可在黄淮海田间进行多点抗性测试,进一步明确其抗性后,再决定是否淘汰。
4 结论
黄淮海玉米主产区与海南三亚玉米南繁基地的玉米小斑病菌在致病力上存在一定差异,黄淮海玉米主产区小斑病菌菌株以弱致病菌群为主,海南三亚以强致病菌群为主。遗传多样性分析结果表明,黄淮海地区与三亚地区的菌株群体具有丰富的遗传多样性,两地菌株整体遗传相似系数较高,遗传距离较近,且两地之间的菌株存在一定的基因交流。三亚地区作为玉米的南繁基地,其自然发病条件下筛选出具有小斑病抗性的玉米品种或抗性亲本材料,对于黄淮海玉米种植区小斑病抗性的逐年提高有一定的影响,其筛选具有应用价值。
[1] 李玥仁, 商鸿生, 胡必德. 陕西省玉米小斑病菌致病性分化研究. 植物保护学报, 1993, 20(1): 90, 96.
LI Y R, SHAGN H S, HU B D. Pathogenic specialization ofin shaanxi province., 1993, 20(1): 90, 96. (in Chinese)
[2] 陆宁海, 吴利民, 郎剑锋, 霍云凤, 石明旺. 新乡地区玉米小斑病菌小种群体结构及致病性分析. 河南科技学院学报(自然科学版), 2016, 44(1): 23-27.
LU N H, WU L M, LANG J F, HUO Y F, SHI M W. Analysis on population structure and pathogenicity differentiation ofin Xinxiang.(, 2016, 44(1): 23-27. (in Chinese)
[3] 代玉立, 甘林, 阮宏椿, 石妞妞, 杜宜新, 陈福如, 杨秀娟. 福建省丙环唑不同敏感性玉米小斑病菌的遗传多样性和致病性. 植物病理学报, 2019, 49(1): 64-74.
DAI Y L, GAN L, RUAN H C, SHI N N, DU Y X, CHEN F R, YANG X J. Genetic diversity and pathogenicity of different propiconazole-sensitive isolates ofin Fujian Province., 2019, 49(1): 64-74. (in Chinese)
[4] TOTH R, SMITH D R. Histopathological changes in susceptible corn inoculated withrace O., 1982, 77(2): 75-82.
[5] 王晓梅, 吕平香, 李莉莉, 见德宝, 杨信东. 玉米小斑病重要流行环节的初步定量研究.Ⅱ病斑产孢、孢子飞散、杀菌剂筛选. 吉林农业大学学报, 2007, 29(2): 128-132.
WANG X M, Lü P X, LI L L, JIAN D B, YANG X D. Preliminary quantitative studies on important epidemic links of. Ⅱ. Lesion sporulation, spore dispersion and fungicide screening., 2007, 29(2): 128-132. (in Chinese)
[6] 董金皋. 农业植物病理学. 2版. 北京: 中国农业出版社, 2013: 98-103.
DONG J G.. Beijing: China Agriculture Press, 2013: 98-103. (in Chinese)
[7] ULLSTRUP A J. The impacts of the Southern corn leaf blight epidemics of 1970-1971., 1972, 10: 37-50.
[8] 张晓慧. 皖北地区玉米小斑病的发生及综合防治. 现代农业科技, 2019(7): 90.
ZHANG X H. Occurrence and integrated control of corn leaf blight in North Anhui., 2019(7): 90. (in Chinese)
[9] 赵真真, 郑训梅, 叶子园, 戴婧, 陈方新, 梅玉云. 安徽省玉米小斑病菌生物学特性的研究. 生物学杂志, 2018, 35(6): 60-63.
ZHAO Z Z, ZHENG X M, YE Z Y, DAI J, CHEN F X, MEI Y Y. Biological characteristics ofin Anhui province., 2018, 35(6): 60-63. (in Chinese)
[10] 代玉立, 甘林, 阮宏椿, 廖蕾, 石妞妞, 杜宜新, 陈福如, 杨秀娟. 福建省玉米小斑病菌的生物学特性研究. 中国农学通报, 2016, 32(31): 131-137.
DAI Y L, GAN L, RUAN H C, LIAO L, SHI N N, DU Y X, CHEN F R, YANG X J. Biological characteristics ofin Fujian Province., 2016, 32(31): 131-137. (in Chinese)
[11] 文静. ISSR分子标记技术在植物病原菌研究中的应用. 安徽农业科学, 2010, 38(36): 20658-20660.
WEN J. Application of ISSR molecular marker technique in plant pathogens research., 2010, 38(36): 20658-20660. (in Chinese)
[12] 李海生. ISSR分子标记技术及其在植物遗传多样性分析中的应用. 生物学通报, 2004, 39(2): 19-21.
LI H S. ISSR molecular marker technology and its application in plant genetic diversity analysis., 2004, 39(2): 19-21. (in Chinese)
[13] 张小飞, 李晓, 崔丽娜, 邹成佳, 李菁, 龙永昌. 我国玉米灰斑病菌遗传多样性的ISSR分析. 植物保护学报, 2015, 42(6): 908-913.
ZHANG X F, LI X, CUI L N, ZOU C J, LI J, LONG Y C. Genetic diversity analysis ofspp. by ISSR in China., 2015, 42(6): 908-913. (in Chinese)
[14] 谷守芹, 范永山, 李坡, 董金皋. 玉米大斑病菌ISSR反应体系的优化和遗传多样性分析. 植物保护学报, 2008, 35(5): 427-432.
GU S Q, FAN Y S, LI P, DONG J G. Optimization of ISSR reaction and genetic diversity analysis of., 2008, 35(5): 427-432. (in Chinese)
[15] 张小飞, 李晓, 崔丽娜, 邹成佳, 杨晓蓉, 向运佳. 玉米圆斑病()遗传多样性ISSR分析. 植物保护, 2015, 41(3): 30-34.
ZHANG X F, LI X, CUI L N, ZOU C J, YANG X R, XIANG Y J. Genetic diversity analysis of corn leaf spot caused bywith ISSR markers., 2015, 41(3): 30-34. (in Chinese)
[16] 郭云燕, 陈茂功, 孙素丽, 武小菲, 江凯, 朱振东, 李洪杰, 何月秋, 王晓鸣. 中国玉米南方锈病病原菌遗传多样性. 中国农业科学, 2013, 46(21): 4523-4533.
GUO Y Y, CHEN M G, SUN S L, WU X F, JIANG K, ZHU Z D, LI H J, HE Y Q, WANG X M. Genetic diversity ofUnderw. in China., 2013, 46(21): 4523-4533. (in Chinese)
[17] 马红霞, 孙华, 郭宁, 张海剑, 石洁, 常佳迎. 禾谷镰孢复合种毒素化学型及遗传多样性分析. 中国农业科学, 2018, 51(1): 82-95.
MA H X, SUN H, GUO N, ZHANG H J, SHI J, CHANG J Y. Analysis of toxigenic chemotype and genetic diversity of thespecies complex., 2018, 51(1): 82-95. (in Chinese)
[18] 常佳迎, 刘树森, 马红霞, 石洁, 郭宁, 张海剑. 黄淮海地区夏玉米弯孢叶斑病菌遗传多样性分析. 中国农业科学, 2019, 52(5): 822-836.
CHANG J Y, LIU S S, MA H X, SHI J, GUO N, ZHANG H J. Genetic diversity analysis ofin summer maize in Huang-Huai-Hai region., 2019, 52(5): 822-836. (in Chinese)
[19] 梅玉云. 安徽省玉米小斑病菌的生物学特性与群体遗传多样性研究[D]. 合肥: 安徽农业大学, 2016.
MEI Y Y. Biological characteristics and population genetic diversities ofin Anhui Province[D]. Hefei: Anhui Agricultural University, 2016. (in Chinese)
[20] 廖蕾. 福建省玉米小斑病菌生物学特性和遗传多样性研究[D]. 福州: 福建农业大学, 2017.
LIAO L. Study on biological characteristics and genetic diversity ofin Fujian Province[D]. Fuzhou: Fujian Agriculture and Forestry University, 2017. (in Chinese)
[21] 常佳迎, 刘莉, 刘树森, 石洁, 郭宁, 张海剑, 刘粤阳. 黄淮海地区夏玉米灰斑病病原菌鉴定及主栽品种抗性分析. 植物病理学报, 2019, 49(6): 808-817.
CHANG J Y, LIU L, LIU S S, SHI J, GUO N, ZHANG H J, LIU Y Y. Identification of causal agent of gray leaf spot and resistance of important hybrids in Huanghuaihai summer maize region., 2019, 49(6): 808-817. (in Chinese)
[22] 张天宇. 介绍一项用于单孢子分离和制片以备显微摄影的显微操作技术. 真菌学报, 1983, 2(3): 197-200.
ZHANG T Y. A technique using micromanipulator for isolating single spores and preparing slide for photomicrogragh., 1983, 2(3): 197-200. (in Chinese)
[23] NEI M. Genetic distance between populations., 1972, 106(949): 283-292.
[24] NEI M. Analysis of gene diversity in subdivided populations., 1973, 70(12): 3321-3323.
[25] 代玉立, 甘林, 滕振勇, 石妞妞, 阮宏椿, 杜宜新, 廖蕾, 陈福如, 杨秀娟. 福建省玉米小斑病菌致病力的分化. 西北农林科技大学学报(自然科学版), 2018, 46(4): 92-97, 106.
DAI Y L, GAN L, TENG Z Y, SHI N N, RUAN H C, DU Y X, LIAO L, CHEN F R, YANG X J. Pathogenicity differentiation ofin Fujian., 2018, 46(4): 92-97, 106. (in Chinese)
[26] WANG M, WANG S Q, MA J, YU C J, GAO J X, CHEN J. Detection ofraces in South China., 2017, 165(10): 681-691.
[27] 王利智, 康志钰, 吴毅歆, 周惠萍, 毛自朝, 何月秋. 云南省玉米小斑病菌生理小种的初步鉴定. 云南大学学报(自然科学版), 2010, 32(3): 352-357.
WANG L Z, KANG Z Y, WU Y X, ZHOU H P, MAO Z C, HE Y Q. Preliminary identification of physiological races ofin Yunnan.(, 2010, 32(3): 352-357. (in Chinese)
[28] 赵聚莹, 蒋晓丽, 贾海民, 李术臣, 石洁, 张海剑. 黄淮海地区玉米小斑病菌生理小种鉴定与评价. 河北农业科学, 2012, 16(9): 47-49.
ZHAO J Y, JIANG X L, JIA H M, LI S C, SHI J, ZHANG H J. Identification and evaluation of physiological races ofin Huanghuaihai region., 2012, 16(9): 47-49. (in Chinese)
[29] 王群. 山东省玉米品种(系)对病害的田间抗性鉴定[D]. 泰安: 山东农业大学, 2018.
WANG Q. Field identification of maize cultivars (lines) resistance to diseases in Shandong Province[D]. Taian: Shandong Agricultural University, 2018. (in Chinese)
[30] 施艳, 燕照玲, 王珂, 赵清爽, 席靖豪, 李冠楠, 刘焱昆, 袁虹霞, 李洪连. 河南省夏玉米品种对6种主要病害的抗性评价. 河南农业科学, 2019, 48(6): 95-98, 105.
SHI Y, YAN Z L, WANG K, ZHAO Q S, XI J H, LI G N, LIU Y K, YUAN H X, LI H L. Evaluation of resistant summer maize varieties against six main diseases in Henan Province., 2019, 48(6): 95-98, 105. (in Chinese)
[31] 陆银萍. 安徽省玉米新品种(系)对玉米主要病害的抗病性研究[D]. 合肥: 安徽农业大学, 2015.
LU Y P. New maize varieties (lines) of resistance to main diseases of maize in Anhui Province[D]. Hefei: Anhui Agricultural University, 2015. (in Chinese)
Pathogenicity and Genetic Diversity ofin Sanya, Hainan and Huang-Huai-Hai Region
CHANG JiaYing, LIU ShuSen, SHI Jie, GUO Ning, ZHANG HaiJian, MA HongXia, YANG Chunfeng
(Plant Protection Institute of Hebei Academy of Agricultural and Forestry Sciences/Key Laboratory of IPM on Crops in Northern Region of North China, Ministry of Agriculture and Rural Affairs/IPM Centre of Hebei Province, Baoding 071000, Hebei)
【Objective】The objective of this study is to clarify the pathogenicity differences, genetic diversity and relationship among thestrains from southern maize breeding base of Sanya, Hainan and Huang-Huai-Hai region, and to provide a theoretical basis for screening resistance of maize varieties under natural disease conditions in the breeding process. It also provides a reference for the popularization and utilization of maize varieties in Huang-Huai-Hai region.【Method】The pathogens were isolated from diseased leaves collected from Sanya, Hainan and Huang-Huai-Hai region, and a total of 61 strains were obtained by identification of morphology and molecular biology. The pathogenicity of 61 strains was identified by an artificial spray inoculation method, and 16 primers with clear, repetitive and polymorphic bands were used for ISSR-PCR. Popgen32 software was used to calculate the genetic distance and genetic similarity among the populations, and NTsys2.10e software was used for cluster analysis and constructing a cluster map of the pathogens in Sanya, Hainan and Huang-Huai-Hai region.【Result】The results of pathogenicity test showed that there was no weak pathogenic strain and 57.1% strong pathogenic strains isolated from Sanya, Hainan, while that of Huang-Huai-Hai region accounted for 47.5% and 17.5%, respectively. The results of genetic diversity analysis showed that the Nei’s gene diversity index (H) was 0.2820 and Shannon’s information index (I) was 0.4197 on the average level of population, indicating that there was a certain genetic variation in the pathogen. The genetic similarity coefficient was 0.9028-0.9618 and the genetic distance was 0.0390-0.1023 among different geographical populations, which indicated that the overall genetic similarity coefficient was relatively high and the genetic distance was close, but there were still different degrees of genetic differentiation among different geographical populations. Among which, the genetic similarity coefficient of strains in Henan and Hebei was the highest (GS=0.9618), and the genetic relationship was the closest; the genetic similarity coefficient of strains in Henan and Sanya was the lowest (GS=0.9028), and the genetic relationship was relatively far. The cluster analysis showed that the strains in Sanya and Huang-Huai-Hai region were obviously divided into two groups when the similarity coefficient was 0.722.【Conclusion】There are some differences inpathogenicity between the main maize producing area of Huang-Huai-Hai region and the southern maize breeding base of Sanya, Hainan. The isolation frequency of the strong pathogenic strains in Sanya was higher than that in Huang-Huai-Hai region. The strain populations in Huang-Huai-Hai region and Sanya region have rich genetic diversity, and genetic diversity has a certain relationship with geographical origin. The genetic relationship of the strains in the adjacent regions was relatively close. However, the genetic similarity coefficient of strains between Sanya region and Huang-Huai-Hai region is relatively high, the genetic relationship is close, and also there are some gene exchanges between the two populations. Therefore, selection of resistant maize varieties or resistant parent materials under natural conditions in southern maize breeding base of Sanya has a certain effect on the yearly increase of resistance to southern corn leaf blight in Huang-Huai-Hai region, which is helpful to reduce the occurrence of disease in Huang-Huai-Hai region.
;southern corn leaf blight; southern maize breeding base; Sanya, Hainan; Huang-Huai-Hai region; pathogenicity; genetic diversity
2019-09-04;
2019-10-23
国家现代农业产业技术体系(CARS-02)
常佳迎,E-mail:cjy198908@163.com。通信作者石洁,E-mail:shij99@163.com
(责任编辑 岳梅)
猜你喜欢
声屏世界(2022年15期)2022-11-08
农业工程学报(2022年5期)2022-06-22
今日农业(2021年21期)2022-01-12
农业与技术(2020年11期)2020-06-22
安徽农学通报(2020年10期)2020-06-12
农民致富之友(2019年29期)2019-10-21
农民致富之友(2019年20期)2019-07-27
校园英语·中旬(2019年5期)2019-07-16
农民致富之友(2019年10期)2019-05-22
安徽农学通报(2016年1期)2016-01-20
