
轮伐期前后不同密度巨桉(Eucalyptus grandis)人工林土壤动物群落结构特征
2020-03-27 09:49:44张阿娟李金金刘志刚张丹桔
生态学报 2020年3期
张阿娟,张 健,李金金,刘志刚,张丹桔
四川农业大学生态林业研究所林业生态工程省级重点验室, 成都 611130
土壤动物不仅是森林土壤生态系统结构和功能的重要组成部分,而且是森林生态系统生物元素的储存者以及生物元素循环和土壤形成与发育的积极参与者[1]。土壤动物与土壤微生物在非生物因子作用下,在维持森林生态系统物质循环和能量流动中起着不可替代的作用,也是指示植被演替、环境干扰及气候变化等的重要生物学指标[2]。我国是世界人工林发展最快、面积最大的国家,总面积已达6933万hm2, 人工林在为区域社会和经济做出巨大贡献的同时,也带来了生物多样性下降、地力衰退等颇具争议的生态环境问题[3]。人工林生态系统在为土壤动物群落提供生存环境的同时,随着其演替及经营措施的改变等方面都会对人工林土壤生物产生深远影响[4]。人工林植被群落的层次结构变化可明显改变土壤水热环境,影响土壤动物群落的生存和繁衍;这些结构和环境的变化还能显著改变土壤动物食物有效性和生存环境,从而影响土壤动物群落结构和功能[4- 7]。因此研究土壤动物群落结构和功能对人工林系统可持续经营和发展具有重要意义。
巨桉(Eucalyptusgrandis),桃金娘科,桉树属植物。由于其生长快、干形好、用途广泛等特点被中国广泛引种栽培,已成为中国南方速生丰产林的战略性树种[8]。四川经过多年引种筛选并营建了大面积的巨桉人工林,轮伐期多在5—7年。本课题组前期在四川省丹棱县研究了一个年龄序列巨桉人工林(1—10年)植物和土壤生物多样性演化特征,发现随林龄增长,土壤动物多样性呈现4年左右降低此后随林龄显著升高的变化趋势,这可能与巨桉人工林随林龄的增加林内和土壤环境的变化有关。前期研究还发现4年和8年可能是巨桉产生化感物质的重要时间节点,4年时巨桉分泌化感物质较多,而进入到土壤中的化感物质可在土壤非生物因子作用下直接或通过影响土壤微生物群落结构等间接影响土壤动物群落[9- 13]。然而,轮伐期前(4年左右)巨桉人工林土壤动物多样性降低的生态作用机制还不清楚。为此,本研究在前期研究基础上,以四川省丹棱县不同密度的轮伐期前(4年)和轮伐期后(8年)巨桉人工林为研究对象,通过手捡、Tullgren及Baermann法分离土壤动物并分析其群落结构特征,旨在探究巨桉人工林土壤动物群落结构动态变化的生态影响因素,为巨桉人工林的科学经营和管理提供理论依据。
1 研究区概况
研究区位于四川省丹棱县杨场镇(102°57′—103°04′ E, 29°55′—29°59′ N),属于亚热带湿润季风气候,海拔550—560 m,年均温17.5 ℃,年降雨量1397 mm,相对湿度82%;土壤为铁铝土性质老冲积黄壤。该地具有大面积处于不同生长阶段(1—10年)和不同密度的巨桉人工林,各林龄林分面积均大于10 hm2。研究地地势平坦(坡度小于5°),通过向当地土地所有者咨询,研究地在巨桉造林前为耕作土壤,其耕作系统以及管理强度在当地具备典型性,耕地轮作制度在巨桉造林前持续多年,因此巨桉人工林样地的土壤基质和原耕作土壤类似;种植巨桉后未经过任何施肥或杂草处理,低林龄林分未经疏伐。该地巨桉人工林轮伐期为5—7年。
2 材料与方法
2.1 样地设置
2017年,选择立地类型相同(轻中壤立地类型)的巨桉人工林,在原有密度基础上通过间伐形成不同密度(D1, 2000; D2, 1600; D3, 1200 株/hm2)的轮伐期前(4年)和轮伐期后(8年)巨桉人工林,每个密度设置3个重复样地,每个1 hm2。2018年4月,在每个样地设置一个20 m×20 m的标准地,样方边界离林缘不小于10 m。采用5点法在各样地采取0—15 cm土样并混合,带回实验室用于测定土壤理化特性,样地基本概况如表1。

表1 4年和8年不同密度巨桉人工林样地概况 (平均值±标准误)
同列不同小写字母表示差异显著(P<0.05); 4,8: 林龄; D1: 密度2000 株/hm2; D2: 密度1600 株/hm2; D3: 密度1200 株/hm2
2.2 样品采集与分离
2018年4月,在每个标准地内随机设置3个样点,彼此间隔10 m以上,样点选择较为平坦、人为活动少且避开斜坡地、洼地、岩石、倒木和大树根。按凋落物层、0—5 cm、5—10 cm和10—15 cm土壤层采集样品。大型土壤动物采样面积为50 cm×50 cm(0.25 m2),用手拣法将收集到的土壤动物放入盛有浓度为75%的酒精容器中杀死,带回实验室在解剖镜下分类计数。采用湿漏斗法(Baermann法)收集小型湿生土壤动物,采用干漏斗法(Tullgren法)收集中小型节肢动物。在采样地各样点收集10 cm×10 cm(0.01 m2)面积的枯落物,再用环刀(R=5 cm,v=100 cm3)自下往上顺次取土,每层取两个土样,用白布袋包好后装入黑布袋,带回实验室分别用Tullgren干漏斗和Baermann湿漏斗分离样品中的土壤动物,其中用于分离湿生的土壤样品采用“四分法”取四分之一进行分离[14]。对分离出的土壤动物进行分类鉴定并计数。
土壤动物的分类鉴定,将捕获所得的土壤动物置于双目解剖镜(Leica, EZ4HD)下观察,主要采用《中国土壤动物检索图鉴》[14]、《中国亚热带土壤动物》[15]、《昆虫分类学》[16]和《幼虫分类学》[17]进行分类鉴定,除湿生土壤动物外,其余物种均鉴定至科水平。
2.3 数据处理与分析
土壤动物类群数量等级划分依据:个体数量大于捕获总量的10.0%以上者为优势类群(+++),占1.0%—10.0%者为常见类群(++),不足1.0%者为稀有类群(+)。
土壤动物功能类群划分依据:本研究将土壤动物功能类群划分为枯食性、捕食性、腐食性、杂食性、植食性、菌食性、尸食性。由于受限线虫在分类过程中并未鉴定至科属,本文参考林英华[20- 21]等将线虫统一划分成杂食性。
群落多样性分析:土壤动物多样性特征主要采用Shannon-Wiener多样性指数(H′)、Margalef丰富度指(D)、Pielou均匀度指数(J)和Simpson优势度指数(C)来计算。式中:Ni为第i类群的个体数;N为群落中所有类群的个体数;Pi=Ni/N,S为类群数。
Shannon-Wiener多样性指数(H′):
Simpson优势度指数(C):
Pielou均匀度指数(E):
E=H′/lnS
Margalef丰富度指数(D):
D=lnS/lnN
采用双因素方差分析(two-way ANOVA)检验林龄和造林密度及其交互作用对土壤动物个体数、类群数、密度和多样性指数的影响;采用单因素方差分析(one-way ANOVA)比较不同密度林地土壤动物个体数、类群数、土壤动物密度和多样性的差异,采用最小显著差异法(LSD)进行两两比较,差异显著性水平为P<0.05;同一密度不同林龄样地土壤动物个体数、类群数、土壤动物密度和多样性指数差异显著性采用独立样本t-测验(Independent-Samplet-test)。DCA分析显示4个轴的梯度最大值小于3,因此使用RDA分析更为合适(置换系数499),用Canoco for Windows 4.5软件中的冗余分析(Redundancy analysis, RDA)对土壤动物主要类群和主要环境因子进行相关性分析。数据的整理与分析主要采用Excel 2010和SPSS 22.0软件,采用Origin 8.5绘制图形。
3 结果与分析
3.1 土壤动物群落组成
本次实验共捕获土壤动物2904只,隶属于4门8纲22目70科。其中大型土壤动物541只,隶属于2门7纲15目30科(表2),优势类群为蚁科和康叭科,占大型动物总捕获量的38.08%;常见类群为跳虫科、鼠妇虫科、疣虫兆科、蜚蠊科等,占大型总捕获量的57.49%,其余4.44%为稀有类群。共捕获中小型土壤动物2363只,隶属于4门8纲16目54科(表3),以大翼甲螨科、等节虫兆科和线虫为优势类群,占中小型动物总捕获量的48.07%;以蚁科、派盾螨科、美绥螨科、康叭科、棘虫兆科等为常见类群,占中小型土壤动物捕获量的41.64%;稀有类群占10.29%。不同密度4年和8年生巨桉人工林土壤动物群落组成不同。在4年生巨桉人工林D1密度,大型土壤动物以蚁科、跳虫科、卵形蛛科、康叭科为优势类群(59.26%),中小型以线虫、寄螨科、等节虫兆科、大翼甲螨科为优势类群(63.01%);D2大型以康叭科、蚁科、疣虫兆科为优势类群(46.74%),中小型以等节虫兆科和线虫为优势类群(40.84%);D3大型以蚁科和鼠妇虫科为优势类群(39.42%),中小型土壤动物以线虫、等节虫兆科、大翼甲螨科为优势类群(52.46%)。8年生巨桉人工林D1密度大型土壤动物以疣虫兆科、蚁科、鼠妇虫科为优势类群(42.42%),中小型以线虫、派盾螨科、大翼甲螨科为优势类群(66.22%);D2大型土壤动物以蚁科和康叭科为优势类群(46.07%),中小型以线虫、派盾螨科、等节虫兆科、大翼甲螨科为优势类群(57.34%);D3大型土壤动物以康叭科、跳虫科、蚁科为优势类群(46.07%),中小型以等节虫兆科、蚁科、线虫为优势类群(50.23%)(表2, 3)。

表2 4年和8年巨桉人工林不同密度大型土壤动物群落组成
—表示无或未采集到, +++为优势类群, ++为常见类群, +为稀有类群; D: 枯食性Debris-feeder′s; S: 腐食性Saprozoic; Pr: 捕食性Predators; O: 杂食性Omnivores; Ph: 植食性Phytophage

表3 4年和8年巨桉人工林不同密度中小型土壤动物群落组成
—表示无或未采集到, +++为优势类群, ++为常见类群, +为稀有类群; F: 菌食性Fungivorous forms; S: 腐食性Saprozoic; Pr: 捕食性Predators; O: 杂食性Omnivores; Ph: 植食性Phytophage; Ca: 尸食性Cadavericoles
3.2 土壤动物群落分布特征
双因素方差分析显示(表4),造林密度对大型土壤动物个体数具有显著影响,对中小型土壤动物类群数有显著影响,同时林龄和密度对中小型土壤动物具有显著的交互作用。林龄和林分密度对大型土壤动物密度有显著影响,但对中小型土壤动物密度无显著影响,二者交互作用对土壤动物密度无显著影响。大型土壤动物类群数及中小型土壤动物个体数、类群数在4年不同密度间均无显著差异;8年生巨桉人工林大型土壤动物个体数及类群数在不同密度无显著差异,中小型土壤动物个体数及类群数随密度降低都显著增大。4年巨桉人工林中小型土壤动物类群数在D1密度显著高于8年,个体数在D1密度以4年高于8年,但其余密度不同林龄之间土壤动物个体数及类群数均表现为8年高于4年(图1)。
大型土壤动物密度在不同土壤层次随林分密度无显著差异。4年生凋落物层中小型土壤动物密度随林分密度降低先显著降低后有所增加;5—10 cm层显著增加,其余层次无显著变化;8年生林地中小型土壤动物密度在凋落物层、0—5及5—10 cm层随林分密度降低均显著增加。在凋落物层,8年巨桉人工林中小型土壤动物密度在D3密度显著高于4年,在其余不同土壤层次,相同密度不同林龄之间土壤动物密度均无显著差异(表5)。

表4 林龄及造林密度对土壤动物作用的双因素方差分析
*P<0.05; **P<0.01;H′: 多样性指数Shannon-Wiener index;C: 优势度指数Simpson index;E: 均匀度指数Pielou index;D: 丰富度指数Margalef index

图1 4年和8年不同密度巨桉人工林土壤动物个体数和类群数Fig.1 The individual number and group number of soil fauna in 4-year old and 8-year old Eucalyptus grandis plantations with different densities 柱状图代表个体数, 折线图代表类群数; *表示同一密度不同林龄间差异显著(P<0.05); 不同小写字母表示同一林龄不同密度间显著差异(P<0.05)

表5 轮伐期前后不同密度巨桉人工林凋落物和各土层土壤动物密度
不同小写字母表示同一林龄不同密度间显著差异(P<0.05);*表示同一密度不同林龄间差异显著
3.3 土壤动物群落多样性指数
双因素方差分析显示,林分密度对大型土壤动物均匀度指数和Margalef丰富度指数有显著影响;对中小型土壤动物Shannon-wiener指数及Margalef丰富度指数有显著影响,林龄及林龄与密度的交互作用不显著(表4)。
4年生巨桉人工林大型土壤动物Margalef丰富度指数随密度降低显著增加,其他多样性指数及中小型土壤动物多样性指数随密度降低无显著变化;8年巨桉人工林大型土壤动物多样性指数随密度降低无显著变化,中小型土壤动物Shannon-wiener指数及Margalef丰富度指数随密度降低显著升高。4年巨桉人工林大型土壤动物Pielou指数在D3密度显著高于8年,4年巨桉人工林中小型土壤动物Shannon-wiener指数在D1密度显著高于8年,在其他密度则无显著变化(表6)。

表6 4年和8年不同密度人工林土壤动物多样性指数
*表示同一密度不同林龄间差异显著(P<0.05); 不同小写字母表示同一林龄不同密度间显著差异(P<0.05)
3.4 土壤动物功能类群特征
研究区域土壤动物营养功能群范围较广,其中大型土壤动物以杂食性为主,占42.42%—75.00%,其次是枯食性(7.35%—21.21%),捕食性(4.41%—19.23%),腐食性(3.45%—15.15%),植食性(4.60%—9.09%)。中小型土壤动物以腐食性为主,占31.30%—52.78%,其次是杂食性(19.78%—40.84%),菌食性(11.03%—24.93%),植食性(4.56%—12.46%),捕食性(0—4.56%),尸食性(0—0.87%)(图2)。

图2 4年和8年不同密度巨桉人工林土壤动物功能类群Fig.2 Functional group of soil fauna in 4-year old and 8-year old Eucalyptus grandis plantations with different densities
3.5 土壤动物与土壤环境因子RDA分析
将4年和8年生不同密度巨桉人工林的主要土壤动物类群与环境因子进行RDA排序。在4年巨桉人工林,土壤含水量与第一排序轴显著正相关,土壤pH、林分密度与第一排序轴呈极显著负相关(图3)。前两个排序轴对物种数据的累计贡献率达54.1%,表明前两个排序轴可以在一定程度上反映不同土壤动物类群与环境因子的关系。8年生巨桉人工林,第一排序轴与林分密度、含水量、土壤pH均呈极显著负相关,与土壤全N显著正相关(P<0.05),第二排序轴与环境因子无显著相关性,前两个排序轴对物种数据的累积贡献率是67.9%,表明前两个排序轴可以在一定程度上反映不同土壤动物类群与环境因子的关系。在RDA排序图中,物种与环境因子线夹角的余弦值可以表征二者的相关性,4年生巨桉人工林的林分密度、土壤pH、含水量对奥甲螨科、长角长虫兆科、寄螨科、隐翅甲科、丽甲螨科等影响较大;8年生巨桉人工林,密度、土壤含水量、pH对线虫、蜚蠊科、康叭科、厉螨科、棘虫兆科等土壤动物影响较大。
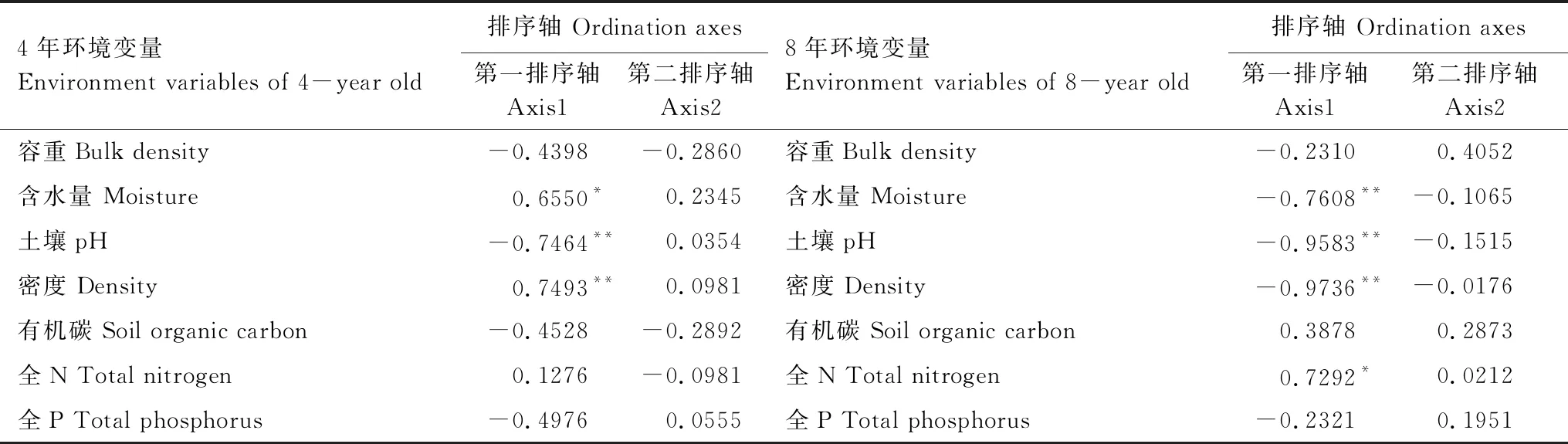
表7 环境变量与排序轴的相关性
*P<0.05; **P<0.01
4 讨论
一般来说,土壤生物的动态变化应该取决于其所需资源的可利用性[18]。巨桉在造林初期,草本植物的迅速发展形成了巨桉人工林的最初净生产力,此后随着林龄的增长,林下植物的生长与死亡成为土壤中C、N和其他营养成分输入的重要来源[18- 19]。因此,由植物提供的可利用性资源决定着土壤的生物多样性。前期研究发现,巨桉人工林植物多样性在1—3年升高,4年左右降低此后随林龄增长而升高;土壤微生物数量和土壤动物多样性呈现1—4年降低,此后随林龄显著升高的变化趋势[9-10]。本研究中,轮伐期前(4年)后(8年)巨桉人工林共捕获土壤动物2904只,分属4门8纲22目70科。土壤动物群落特征随林龄和密度具有一定差异。

图3 4年和8年不同密度巨桉人工林土壤动物与环境因子RDA二维排序图Fig.3 A two-dimensional graph of RDA ordination for soil fauna and the environmental factors in 4-year old and 8-year old Eucalyptus grandis plantations with different densitiesD: 密度Density; MC: 含水量Moisture content; pH: 土壤pH; BD: 容重Bulk density; TP: 全磷Total phosphorus; SOC: 有机碳Soil organic carbon; TN: 全氮Total nitrogen; Stig: 长须螨科Stigmaeidae; Orch: 长角长姚科Orchesellidae; Nean: 疣虫兆科Neanuridae; Stap: 隐翅甲科Staphylinidae; Form:蚁科Formicidae; Ench: 线蚓科Enchyiraeidae; Nema: 线虫Nemata; Term: 螱科Termitidae; Podu: 跳虫科Poduridae; Proc: 鼠妇虫科Procellionidae; Parh: 派盾螨科Parholaspididae; Amer: 美绥螨科Ameroseiidae; Lohm: 罗甲螨科Lohmannidae; Roti: 轮虫Rotifera; Oono: 卵形蛛科Oonopidae; Liac: 丽甲螨科Liacaridae; Lacc: 厉螨科Lacclapidae; Camp: 康叭科Campodeidae; Phth: 卷甲螨科Phthiracaridae; Scia: 尖眼蕈蚊科Sciaridae; Para: 寄螨科Parasitidae; Onyc: 棘虫兆科Onychiuridae; Anis: 肥螋科Anisolabididae; Blat: 蜚蠊科Blattidae; Isot: 等节虫兆科Isotomidae; Galu: 大翼甲螨科Galumnidae; Oppi: 奥甲螨科Oppiidae
总体来看,巨桉人工林造林密度显著影响了大型土壤动物个体数和土壤动物多样性指数,林龄显著影响了大型土壤动物密度,且这两个因素具有一定的交互作用;8年生林地土壤动物个体及类群数显著高于4年。在土壤生态系统中,土壤动物与微生物一起在非生物因子的共同作用下影响地上植物的生长[20]。而植被的组成及其结构变化常常决定着生态系统中其他成分的组成和结构特征[21]。大量研究表明,土壤动物群落结构与植被状况密切相关,植物群落变化过程中土壤动物群落也发生着改变。因此,土壤动物群落组成和结构对生态系统植被结构和演替具有重要的指示作用[1-2]。在人工林生态系统中,由于生长阶段或林分密度的不同,改变了系统的植被组成结构,相应地引起林内微环境的变化,包括光照、空气湿度、土壤含水量、地温的日变化和空间分布规律,以及降水的再分配等[21]。张柳桦等研究发现林分密度对桉树的生物量影响显著[22]。林分密度的降低使巨桉人工林林内光照增强,进而导致土壤温度的升高,加快了土壤中有机质的分解,使土壤速效养分增加,从而提高了人工林土壤肥力[22- 24]。土壤环境因子的变化引起土壤动物个体数及类群分布差异,决定了林分密度的变化也会影响到土壤生态系统中土壤动物的群落结构组成[25- 26]。本研究中在D1密度中小型土壤动物类群数表现为4年显著高于8年,个体数在D1密度为4年高于8年,但其余密度类群数及个体数均以8年较高,且总体来看8年巨桉人工林土壤动物个体数及类群数较4年较高。巨桉在轮伐期前4年左右生长迅速,林分对土壤养分和水分竞争较大,林下植被较少,但巨桉的凋落物量较大,此时巨桉可能通过挥发、雨雾淋溶、凋落物分解和此时较为旺盛的根分泌释放较多的化感物质,这些都会对土壤动物有一定的毒害作用[1,12,27],这些因素都会导致4年时林分群落结构不稳定从而使得土壤动物群落尤其是中小型土壤动物个体、类群及多样性等方面波动较为明显;而随着林龄的增长,林下植被群落结构逐渐稳定,由于巨桉树皮和叶片凋落使得林分此时的光照水平得到改善,促进了土壤微生物群落的发展,加快了土壤有机质分解,这些都有利于提高土壤动物群落的多样性和稳定性[9,28- 31]。
本研究中,轮伐期前后不同密度巨桉人工林环境因子与土壤动物优势类群的相关性分析显示,轮伐期前(4年)林分密度、土壤含水量和pH值为对土壤动物的主要环境作用因子,奥甲螨科、长角长虫兆科、寄螨科、隐翅甲科、丽甲螨科等受影响较大;轮伐期后(8年)林分密度、土壤含水量和pH值、土壤N含量是主要作用因子,对线虫、蜚蠊科、康叭科、厉螨科、棘虫兆科等土壤动物影响较大。说明轮伐期前这些环境因子主要对中小型腐食性土壤动物影响较大,而轮伐期后这些环境因子主要影响杂食性土壤动物,在8D3样地中,由于其林分密度最低,有机质、全氮含量均较高,该地土壤动物个体数、类群数也高于其他样地,这可能是较低的密度使林内光照增强导致土温升高,促进了土壤微生物群落发展,加速了有机质的分解,有利于中小型杂食性、腐食性和菌食性土壤动物的生存[27],增加了一些对环境敏感的稀有物种的生存能力,出现了诸如原铗叭科、奇蝽科、管巢蛛科、足角螨科等的稀有物种。研究表明较低林分密度更有利于保持土壤动物群落多样性和稳定性,而轮伐期前后土壤动物均受林分密度、土壤含水量和pH影响,说明林分密度的改变使植被群落结构发生变化从而对土壤动物群落结构有显著影响。
土壤动物多样性反映土壤中食物资源和生境的多样性,多样性指数比个体数及类群数更能体现群落的结构,在理论和实践中具有重要意义[32- 34]。不同密度4年和8年生巨桉人工林土壤动物多样性具有显著差异,这种差异与生境的异质性紧密相关。一般来说,生境的空间异质性越高,可提供的小生境及小气候条件越丰富,也就意味着更高的生物多样性[35]。本研究中双因素方差分析显示林分密度对大型土壤动物和中小型土壤动物都有显著影响,并且进一步LSD多重比较表明4年生巨桉人工林大型土壤动物Margalef丰富度指数随密度降低显著增加;8年巨桉人工林中小型土壤动物Shannon-wiener指数及Margalef丰富度指数随密度降低显著升高。随着林分密度的降低,土壤温度升高,土壤微生物数量和活性有所升高,能为不同功能群的土壤动物提供多样化的栖息环境和食物源,因而较低林分密度的巨桉人工林土壤动物多样性指数较高。
营养结构是物种群落类群间和物种间相互关系的外在体现,其结构决定着群落功能的发挥[36]。本研究中大型土壤动物以杂食性为主,枯食性和捕食性次之,中小型土壤动物以腐食性为主,杂食性和菌食性次之。一般认为中小型土壤动物对环境变化的反应较为敏感,大型土壤动物移动力较强,营养结构更为复杂,受环境因子影响反应不如中小型土壤动物敏感[9,37- 39]。本研究中,中小型土壤动物以大翼甲螨科、等节虫兆科和线虫为优势类群,占中小型动物总捕获量的48.07%。大翼甲螨科是甲螨亚目常见的腐食性土壤动物,以取食菌类、藻类、腐烂的植物为生,对有机质的分解具有重要作用。等节虫兆科为弹尾目菌食性昆虫,通称跳虫,主要生活在有机质较高的土壤表层,多以真菌为生,是生态系统物质分解与转化的再加工者。土壤线虫种类丰富,数量繁多,分布广泛,是土壤动物中十分重要的杂食性类群,它们在土壤生态系统中占有多个营养级,与其他土壤生物形成复杂的食物链网,在维持生态系统稳定、促进物质循环和能量流动方面起着不容忽视的作用[1,40]。杂食性和腐食性功能群的生态功能是分解枯落物、促进土壤团粒结构的形成、取食微生物以调控整个生态系统物质循环和能量流动的速率等,杂食性土壤动物生态位广泛,生存适应能力强,腐食性功能群常被作为土壤肥力高低的指标之一[1,41- 42]。
5 结论
本研究中,不同密度轮伐期前后即4年和8年生巨桉人工林土壤动物群落特征随林龄和林分密度的改变具有差异。轮伐期后较轮伐期前土壤动物个体数、类群数及多样性指数提高,土壤动物营养结构更为合理,这与轮伐期前后巨桉的生长及林内环境特征如土壤含水量、pH和土壤N含量等有关,轮伐期前后巨桉人工林土壤动物以杂食性和腐食性为主。随着林分密度的降低,土壤动物群落多样性和丰富度均显著提高。建议适当延长巨桉人工林轮伐期并降低林分密度可以改善土壤动物生境、多样性及群落结构的稳定性,将有助于巨桉人工林的科学经营和管理。
猜你喜欢
当代水产(2021年10期)2022-01-12 06:21:02
防护林科技(2020年10期)2020-12-22 11:46:42
江苏农业科学(2019年5期)2019-09-02 14:01:46
上海包装(2019年2期)2019-05-20 09:10:56
广东农业科学(2017年5期)2017-08-29 10:37:31
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
广东石油化工学院学报(2016年3期)2016-05-17 05:16:24
中国粮油学报(2016年5期)2016-01-23 02:44:49
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
